Эта статья опубликована под лицензией Creative Commons и не автором статьи. Поэтому если вы найдете какие-либо неточности, вы можете исправить их, обновив статью.

Генотипическая обусловленность пигментного состава хвои плюсовых деревьев ели европейской



Бессчетнова Н.Н.
П.В. Ершов
В.П. Бессчетное
Опубликована Янв. 1, 2019
Последнее обновление статьи Сен. 18, 2022
Эта статья опубликована под лицензией
")



Аннотация
Изучен пигментный состав 1- и 2-летней хвои плюсовых деревьев (ортетов) ели европейской из состава архива клонов № 3 Государственного бюджетного учреждения Нижегородской области «Семеновский спецсемлесхоз». где они представлены привитыми растениями (клонами или раметами). Цель исследований - выявление факта наличия генотипически обусловленной специфики плюсовых деревьев ели европейской по составу пигментов, участвующих в фотосинтезе, и установление масштабов ее проявления в 1- и 2-летней хвое. Все учетные растения имели одинаковые площади питания и схемы размещения, получали единый перечень лесоводственных и агротехнических уходов. Отбор образцов хвои проведен одновременно с учетом ее возраста и принадлежности к ортетам и раметам. Анализируемые показатели: содержание хлорофилла а. хлорофилла Ь. каротиноидов, их суммарное количество, соотношение между ними. В эксперименте использован спектрофотометрический анализ как наиболее эффективный способ количественного выявления важнейших пластидных пигментов. Экстракционный агент - 96 %-й этанол. Максимумы поглощения при количественной фиксации оптической плотности вытяжек соответствовали следующим длинам волн: 663 нм - для хлорофилла а; 645 нм - для хлорофилла Ь; 440 нм - для недифференцированных каротиноидов. Для статистической обработки привлечен одно- и двухфакторный дисперсионный анализ. Установлены заметные различия пигментного состава листового аппарата плюсовых деревьев ели европейской при выращивании на выровненном экофоне. Однофакторный дисперсионный анализ подтвердил их существенность для 1- и 2-летней хвои по всем задействованным в опыте признакам. Влияние специфики генотипов плюсовых деревьев, эффект которого признается в качестве коэффициента наследуемости в широком смысле, достаточно велико и достоверно по всем рассматриваемым признакам. Раздельное влияние специфики генотипов плюсовых деревьев и различий в возрасте их хвои на формирование фенотипической изменчивости зафиксировано в двухфакторном дисперсионном анализе. Влияние возраста хвои на формирование различий в содержании отдельных пигментов и их групп достоверно, но невелико и в целом составило от 7.10±0.16 % (содержание каротиноидов) до 9.40±0.15 % (суммарное содержание хлорофилла). Доля дисперсии, связанной со спецификой генотипов плюсовых деревьев, в большинстве случаев значительна: до 48.87 %. Взаимодействие организованных факторов чаще вызывает достоверный эффект, однако доля этого организованного фактора в структуре общей фенотипической дисперсии практически
никогда не преобладает. Наличие остаточной дисперсии обусловлено неизбежной пестротой экологического фона.
Ключевые слова
Ель европейская, генотипическая обусловленность, каротиноиды, хлорофилл b, пигментный состав, плюсовые деревья, архив клонов, хлорофилл a, хвоя
Введение
Одним из главных условий устойчивого развития отечественного лесного хозяйства является наличие эффективно функционирующей постоянной лесосеменной базы [7]. В качестве ключевого аспекта ее формирования в России и в других государствах рассматриваются плюсовые деревья основных лесообразующих пород [3, 14, 15, 26, 34]. В их числе ель европейская по праву занимает лидирующие позиции [14, 15, 22]. Обладая хозяйственно важными признаками в сочетании с выраженной устойчивостью и экологической пластичностью, она выступает предметом многочисленных и разноплановых исследований отечественных [4, 5, 8, 13, 16] и зарубежных [22, 25, 40] ученых. Немалая часть этих работ посвящена всестороннему изучению листового аппарата.
Пигментный состав хвои вполне справедливо относят к важнейшим биологическим характеристикам древесных видов, связанным с режимом фотосинтеза, продуктивностью и адаптивностью [12]. Обоснованно признается его защитная роль: присутствие хлорофилла и каротиноидов усиливает резистентность растений [10, 37]. Динамику пигментного состава считают адаптивной реакцией хвойных, в том числе и ели европейской, на условия освещенности [20, 35, 36, 38, 39, 41]. Известно, что в направлении с юга на север в географическом масштабе более устойчивы растения с повышенной пигментацией листового аппарата [10]. Обнаружив преобладание сильно пигментированных форм в центрах формообразования, Н И. Вавилов рассматривал это явление как защитную реакцию растений на действие мутагенных факторов [6]. Сдвиги в содержании хлорофилла а, хлорофилла Ъ, их суммарном количестве и концентрации каротиноидов являются реакцией на изменение концентрации углекислого газа в атмосфере [28, 33]. Известна зависимость содержания зеленых пигментов хвои от температуры почвы и других почвенных характеристик [40]. Сообщается о фотопериодической реакции, которая выражается в изменении содержания пигментов [40], реакции на содержание озона [23], на загрязнение [27]. Вместе с тем у псевдотсуги Мензиса существенных изменений в концентрации пигментов под влиянием ультрафиолетовых лучей не отмечено [19]. Пигментный состав хвои в значительной мере контролируется генотипом и является в значительной степени наследуемым показателем [2, 3], что подтверждается испытаниями потомств от контролируемых скрещиваний [33] и генетическими исследованиями [32]. Другие специалисты [14], напротив, считают, что степень генотипической детерминации признаков пигментного состава хвойных невысока. Отсутствие единого мнения о наследственной специфике указанных признаков и свойств у древесных растений служит достаточным основанием для продолжения работ в указанном направлении, в частности в отношении ели европейской.
Цель исследований - выявление факта наличия генотипически обусловленной специфики плюсовых деревьев ели европейской по составу пигментов, участвующих в фотосинтезе, и установление масштабов ее проявления в 1- и 2-летней хвое.
Объекты и методы исследования
Объектами исследований выступали плюсовые деревья ели европейской, выделенные в естественных насаждениях Нижегородской области согласно действовавшим на момент отбора инструкциям и нормативам. Они входили в состав природных популяций данного вида, типичных для лесорастительного района хвойно-широколиственных (смешанных) лесов европейской части Российской Федерации. Их вегетативные репродукции зарезервированы в архиве клонов № 3 Государственного бюджетного учреждения Нижегородской области «Семеновский спецсемлесхоз». Возраст привитых растений одинаков и согласно паспорту архива клонов составляет 34 года. Все клоны (раметы) каждого из изученных плюсовых деревьев (ортетов) в пределах указанного архива были размещены по единой посадочной схеме с одинаковой площадью питания. Режим содержания архива предполагал общий комплекс агротехнических и лесоводственных мероприятий, прочие условия произрастания также были идентичными. Следует учитывать, что существующий регламент создания и содержания таких объектов единого генетике- селекционного комплекса направлен на механическое выравнивание поверхности участка, отводимого под них, на сглаживание расхождений в его локальных почвенных характеристиках. Такой организационный подход не создавал ни одному учетному дереву преимуществ в доступе к ресурсам роста и развития и обеспечивал принцип единственного логического различия. Возраст хвои выступал одним из организованных факторов дисперсионного анализа. В соответствии с этим все остальные эффекты, связанные с возможным возникновением временной изменчивости, нивелировались или исключались полностью. Это достигалось одновременным выполнением отбора биологических образцов для анализа и одинаковыми сроками проведения всех его этапов. Формирование выборочных совокупностей и включение в их состав первичных единиц выборки выполнены в общепринятом порядке [9, 11]. Реализованы полевой стационарный и лабораторный опыты. Их организационная схема позволила сформировать двухфакторные равномерные и пропорциональные дисперсионные комплексы, в которых каждый ортет был представлен 3 раметами. С периферии среднего яруса хорошо освещенного участка из кроны срезали по 5 ветвей, из средней части которых отделяли хвою для последующей экстракции пигментов. В итоге по каждому признаку получено 315 первичных единиц выборки. Поскольку присутствие хлорофилла и каротиноидов в хвое коррелирует с устойчивостью растений к лимитирующим факторам среды [1, 12, 37], проведение анализов установлено на декабрь 2016 г.
Определение содержания пигментов, участвующих в фотосинтезе, реализовано традиционными методами в ходе камерального этапа исследований [17, 18, 29-31, 40, 42]. Вполне результативным и достаточно точным методом исследования пигментного состава листового аппарата признан спектрофотометрический анализ [17, 18, 21, 24, 31, 42]. С его помощью удается оценить концентрацию пигментов по оптической плотности вытяжки из измельченной хвои в оптически чистом ацетоне или в 96 %-м этаноле. В работе был задействован электроспектрофотометр Grating 722, при этом максимумы поглощения при количественной фиксации оптической плотности вытяжек соответствовали следующим длинам волн: 663 нм - для хлорофилла щ 645 нм - для хлорофилла ö; 440 нм - для недифференцированных каротиноидов. Известно, что положение максимума поглощения в спектральном анализе сдвигается в ту или иную сторону при использовании разных экстрагентов [17, 18, 21, 31, 42]. Это учитывалось в эксперименте. Содержание анализируемых пигментов
рассчитывали по уравнениям Ветштейна и Хольма [17, 42], при этом для нахождения концентрации, выраженной в миллиграммах на грамм, по каждой навеске определяли содержание абсолютно сухого вещества. В построении алгоритмов для указанных расчетов учитывали известные методические приемы [28-31]. Анализировали содержание и соотношения хлорофилла а, хлорофилла Ъ, каротиноидов, их доли и суммарное количество. Работу строили на платформе комплексности многопараметрических исследований [2, 3], целесообразность которых общепризнана [28-31, 40]. Первичную лесоводствен- ную информацию обрабатывали с использованием известных статистических схем [9, 11].
Результаты исследования и их обсуждение
Установлены заметные различия пигментного состава листового аппарата плюсовых деревьев ели европейской при выращивании на выровненном экофоне. Одно факторный дисперсионный анализ подтвердил их существенность для 1- и 2-летней хвои по всем задействованным в опыте признакам (табл. 1).
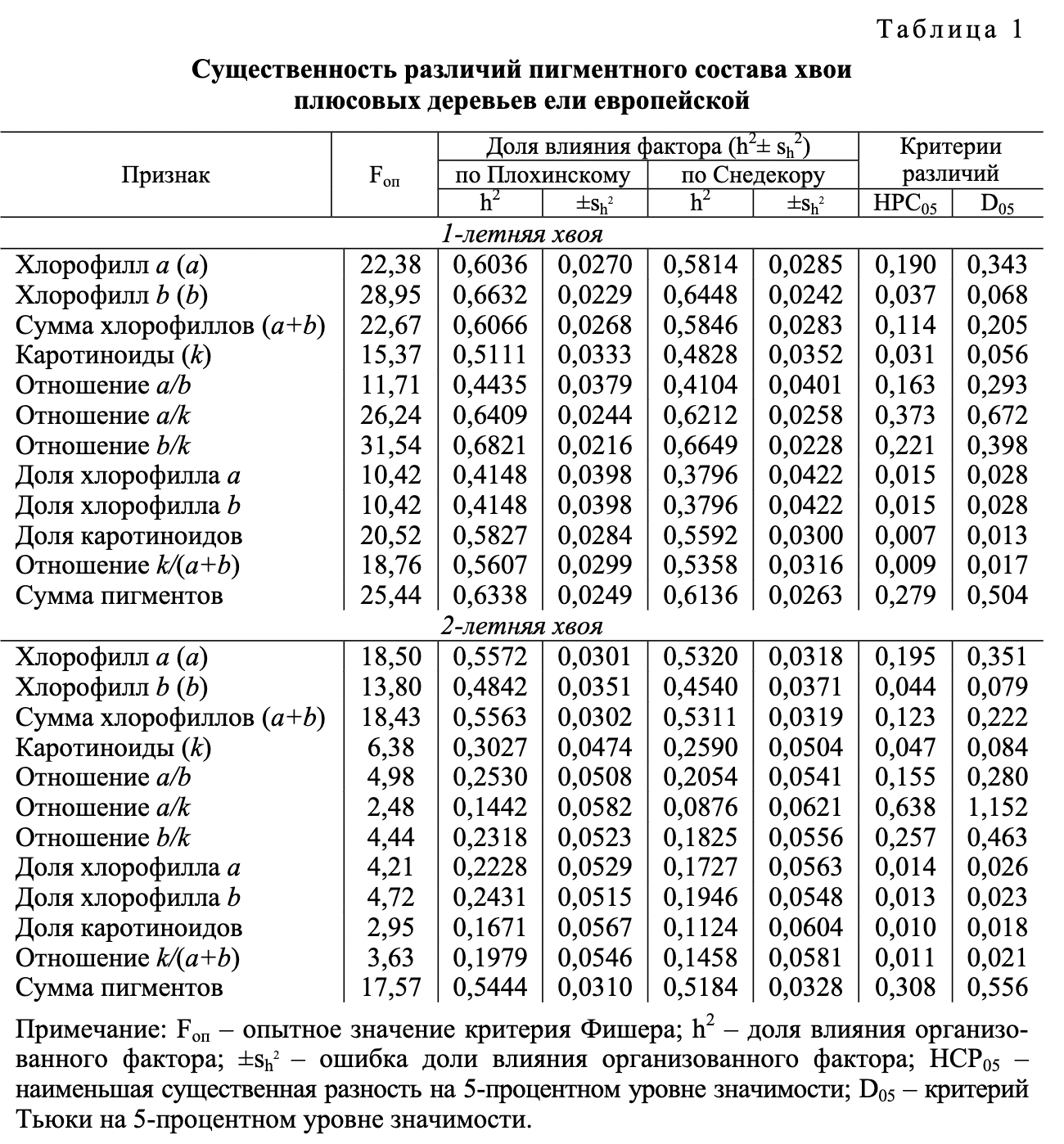
Для хвои всех возрастов опытные значения критерия Фишера (табл. 1) преодолели критический порог как на 5-, так и на 1-процентном уровне значимости. В большой мере это обусловлено достаточным числом первичных единиц выборки, введенных в каждый дисперсионный комплекс. Влияние специфики генотипов плюсовых деревьев, эффект которой рассматривается как коэффициент наследуемости в широком смысле, по всем анализируемым показателям достоверно и достаточно велико. В частности, для 1-летней хвои в вычислениях по алгоритму Плохинского содержание хлорофилла а составило 60,36±2,70 %, для хлорофилла b оно было еще выше - 66,32±2,29 %. Максимум (68,21±2,16 %) достигнут для отношения содержания хлорофилла b к содержанию каротиноидов. Расчетный метод Сне декора адекватно показал близкие по значениям итоги вычислений.
Для 2-летней хвои указанный эффект оказался несколько ниже, но вполне сопоставим по значениям: по содержанию хлорофилла-п - 55,72±3,01 %, хлорофилла-0 - 48,42±3,51 %. Достаточно высокие результаты в этом случае отмечены для суммарного содержания хлорофилла (55,63±3,02 %) и общей суммы пигментов (54,44±3,10 %). Вместе с тем оценки наследственной обусловленности наличия каротиноидов в 2-летней хвое были ниже, чем в 1-летней хвое. Снизились результаты и других признаков. Критерии существенности различий (НСРо5 и Dos) определяют уровень, достигнув которого, разница между средними значениями характеристик плюсовых деревьев признается существенной, т. е. обусловленной преимущественно их наследственными особенностями.
Существенность различий плюсовых деревьев ели европейской по пигментному составу, подтвержденная для 1- и 2-летней хвои, определила возможность выявления эффекта индивидуального влияния каждого из организованных факторов на общую фенотипическую дисперсию. В частности учтено влияние различий между самими плюсовыми деревьями и различий в возрасте их хвои отдельно по каждому из этих факторов (табл. 2).
Практически во всех случаях учета организованные факторы генерируют дисперсию, уровень проявления которой соответствует существенным различиям между элементами дисперсионного комплекса. Опытные критерии Фишера оказались значительно больше соответствующих нормативных значений на 5-процентном уровне значимости. Исключением явились оценки по разновозрастной хвое для отношений содержания хлорофилла а к содержанию каротиноидов и содержания хлорофилла Ъ к содержанию каротиноидов. Это указывает на индифферентность баланса различных форм хлорофилла и каротинодов в отношении увеличения возраста хвои. Выраженная хронографическая стабильность указанных характеристик плюсовых деревьев при достаточно высокой их генотипической обусловленности (22,48^2,64 % и 30,59±2,36 % соответственно) позволяет признать значительную информативность и большое идентификационное значение соотношений между двумя формами хлорофилла и каротиноидами. В соответствии с этим указанные признаки можно рекомендовать к использованию для оценки (при определении) специфичности плюсовых деревьев в процессе селекционной инвентаризации объектов постоянной лесосеменной базы и единого генетикоселекционного комплекса, а также при выработке рекомендаций по их включению в состав лесосеменных плантаций.
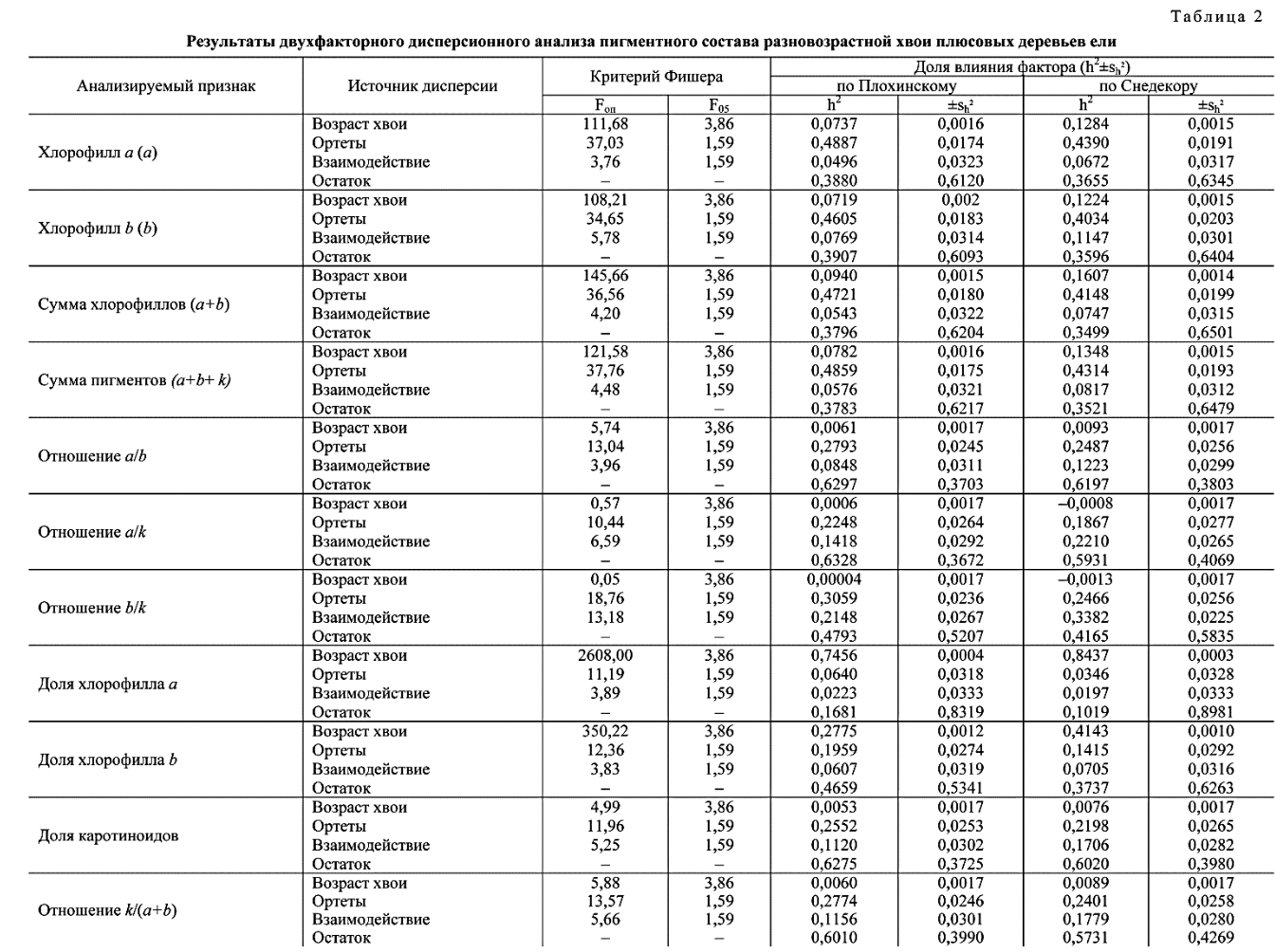
Влияние возраста хвои на формирование различий в содержании отдельных пигментов и их групп (табл. 2) достоверно, но невелико и в целом составило от 7,10±0,16 % (содержание каротиноидов) до 9,40±0,15 % (суммарное содержание двух форм хлорофилла). Оценки мало различаются, и наибольшая из них превосходит наименьшую в 1,32 раза, при том что общий размах значений в вариантах с подтвержденной существенностью различий оценивается соотношением 124,27. Изменение с возрастом доли содержания в хвое хлорофилла а более заметно по сравнению с другими характеристиками пигментного состава. В частности, влияние фактора «возраст хвои» достигает
74,56±0,04 % и может быть связано с ощутимым повышением содержания данного пигмента в 2-летней хвое. Отчетливо выражена реакция на увеличение возраста хвои в содержании хлорофилла Ь : 27,75±0,12 %.
Влияние различий между плюсовыми деревьями на общий фон изменчивости анализируемых характеристик пигментного состава их листового аппарата занимает значительную часть в большинстве случаев. Величина этого эффекта составляет от 23,62±2,60 % (содержание каротиноидов) до 48,87±1,74 % (содержание хлорофилла а) и 48,59±1,75 % (сумма пигментов), что зафиксировано в отношении содержания отдельных пигментов и их суммарного количества. Характеристики соотношения исследуемых пластидных пигментов в большей мере связаны с наследственной спецификой ортетов и меньше зависят от возраста хвои. Вместе с тем наследственная обусловленность различий между плюсовыми деревьями в этом случае выражена в несколько меньшей степени, чем для оценок содержания пигментов.
Взаимодействие организованных факторов чаще вызывает достоверный эффект, однако доля этого организованного фактора в структуре общей фенотипической дисперсии практически никогда не преобладает. Остаточная дисперсия, наличие которой, как правило, связывают с пестротой параметров среды на объектах постоянной лесосеменной базы и единого генетике- селекционного комплекса, к которым относится исследованный архив клонов, имеет неоднозначные оценки. При этом в ряде рассматриваемых признаков она доминирует, что зафиксировано в отношении содержания хлорофилла-п к содержанию каротиноидов (62,97 %) и содержания каротиноидов к суммарному содержанию хлорофилла (60,10 %). В целом указанные результаты вполне адекватны сведениям, полученным в процессе однофакторного дисперсионного анализа как по 1-летней, так и по 2-летней хвое.
Заключение
Плюсовые деревья ели европейской существенно различаются по широкому спектру показателей пигментного состава 1- и 2-летней хвои. Степень наследственной обусловленности проявлений разных признаков неодинакова. Оценки содержания хлорофилла а, хлорофилла Ъ и их суммарного количества, содержания каротиноидов и общей суммы пигментов тесно связаны с генотипом исследованных клонов. Показатели соотношения между пластидными пигментами в целом имеют меньшую степень генотипической обусловленности по сравнению с количественными оценками их содержания. Оценки отношения хлорофилла а и хлорофилла Ъ к содержанию каротиноидов весьма стабильны в онтогенетическом плане, что свидетельствует об их большом идентификационном значении и указывает на целесообразность применения при ревизии чистоты ассортиментного состава лесосеменных плантаций и архивов клонов.
СПИСОК ЛИТЕРАТУРЫ
- Барская Е.П. Изменения хлоропластов и вызревание побегов в связи с морозоустойчивостью древесных растений. М.: Наука. 1967. 223 с.
- Бессчетнова Н.Н. Многомерная оценка плюсовых деревьев сосны обыкновенной по показателям пигментного состава хвои // Вести. ПГТУ Сер.: Лес. Экология. Природопользование. 2013. № 1. С. 5-13.
- Бессчетнова Н.Н., Бессчетное В.П. Сосна обыкновенная (Pinus sylvestris). Морфометрия и физиология хвои плюсовых деревьев. Н. Новгород: Нижегород. ГСХА. 2014. 368 с.
- Бессчетнова, Н.Н., Кулькова А.В. Сравнительная оценка представителей рода ель (Picea) по содержанию жиров в тканях годичных побегов // Научные и инновационные разработки молодых ученых-аграриев: сб. тр. молодых ученых ФГБОУ ВПО Нижегородская ГСХА за 2014-2015 гг. Н. Новгород: Нижегород. ГСХА. 2015. С. 53-58.
- Бессчетнова Н.Н., Бессчетное В.П., Кулькова А.В., АІишукова II.В. Содержание крахмала в тканях побегов разных видов ели (Picea Dietr.) в условиях интродукции // Леей. жури. 2017. № 4. С. 57-68. (Изв. высш. учеб, заведений). DOI: 10.17238/issn0536-1036.2017.4.57
- Вавилов Н.П. Учение о происхождении культурных растений после Дарвина // Избр. тр. в 2 т. Т. 1. Л.: Наука. 1967. С. 303-327.
- Государственная программа Российской Федерации «Развитие лесного хозяйства» на 2013-2020 годы: утв. распоряжением Правительства Российской Федерации от 28 дек. 2012 г. № 2593-р // Собр. законодательства Российской Федерации. 2013. № 2. Ст. 124. Режим доступа: http://www.nbchr.ru/PDF/042_oos.pdf (дата обращения: 17.09.2013).
- Дерюгин А.А., Рубцов М.В. Динамика состояния популяции ели под пологом березняков южной тайги Русской равнины // Леей. жури. 2016. № 2. С. 47-58. (Изв. высш. учеб, заведений). DOI: 10.17238/issn0536-1036.2016.2.47
- Доспехов БА. Методика полевого опыта (С основами статистической обработки результатов исследований). Изд. 5-е. доп. и перераб. М.: Агропромиздат. 1985. 351с.
- Моссэ II.Б., Молофей В.П., Кострова Л.Н. Развитие идеи Н И. Вавилова о защитной роли пигментов в генетических экспериментах // Селекция, генетические ресурсы и сохранение генофонда лесных древесных растений (Вавиловские чтения): сб. науч. тр. Гомель. 2003. Вып. 59. С. 220-223.
- Никитин КБ., Швиденко А.З. Методы и техника обработки лесоводствен- ной информации М.: Леси, пром-сть. 1978. 272 с.
- Озолина ПА., Мочалкин А.II. Роль пигментов в защитно-приспособительных реакциях растений//Изв. АН СССР. Сер. Биол. 1972. № 1. С. 96-102.
- Пеккоев А.Н. Качество древесины культур ели при ускоренном лесовыра- щивании // Леси. жури. 2016. № 1. С. 89-99. (Изв. высш. учеб, заведений). DOI: 10.17238/issn0536-1036.2016.1.89
- Роне В.AI Исследование и проверка наследственности плюсовых деревьев // Леси, селекция. М.: Леси, пром-сть. 1972. С. 180-199.
- Роне В.AI. Межсемейный и клоповый отбор ели обыкновенной // Генетические исследования древесных в Латвийской ССР. Рига: Зинатне. 1975. С. 34-44.
- Сунгурова Н.Р., Коновалов В.Н. Ассимиляционный аппарат ели в культурах на долгомошной вырубке // Леси. жури. 2016. № 2. С. 31-40. DOI: 10.17238/issn0536- 1036.2016.2.31
- Третьяков Н.Н., Карнаухова Т.В., Паничкин ЛА. и др. Практикум по физиологии растений: учеб, пособие для студентов высш. учеб, заведений / под общ. ред. Н.Н. Третьякова. 3-е изд., перераб. и доп. М.: Агропромиздат. 1990. 271 с.
- Шлык АА. Определение хлорофиллов и каротиноидов в экстрактах зеленых листьев//Биохимические методы в физиологии растений. М.: Наука. 1971. С. 154-170.
- Bassman J.H., Edwards G.E., Robberecht R. Long-Tenn Exposure to Enhanced UV-B Radiation is not Detrimental to Growth and Photosynthesis in Douglas-Fir // New Phytologist. Vol. 154. iss. 1. Pp. 107-120. D01:10.1046/j.l469-8137.2002.00354.x
- Bouvier /•’., Backhaus R.A., Camara B. Induction and Control of Chloroplast- Specific Carotenoid Genes by Oxidative Stress // Journal of Biological Chemistry. Vol. 273. no. 46. Pp. 30651-30659. DOI: 10.1074/jbc.273.46.30651
- Dere .S', Günes T„ Sivaci R. Spectrophotometric Determination of Chlorophyll - A. В and Total Carotenoid Contents of Some Algae Species Using Different Solvents // Turkish Journal of Botany. Vol. 22. no. 1. Pp. 13-17.
- Eerikäinen К., Va Ikon en S., Saksa T. Survival and Height Growth of Small Trees in Uneven-Aged Picea abies Stands in Southern Finland I I Forest Ecosystems. 2014. Vol. 1(5). Pp. 1-10. DOI: 10.1186/2197-5620-1-5
- Elvira .S', Alonso R., Castillo F.J, Gimeno B.S. On the Response of Pigments and Antioxidants of Pi mis halepensis Seedlings to Mediterranean Climatic Factors and Long- Term Ozone Exposure // New Phytologist. Vol. 138. iss. 3. Pp. 419-432. DOI: 10.1046/j. 1469-8137.1998.00136.x
- Gitelson A.A., Buschmann C., Lichtenthaler H.K. The Chlorophyll Fluorescence Ratio F735/F7oo as an Accurate Measure of the Chlorophyll Content in Plants 11 Remote Sensing of Environment. Vol. 69. iss. 3. Pp. 296-302. DOI: 10.1016/S0034- 4257(99)00023-1
- Haag ('., Stober /•’., Lichtenthaler H.K. Pigment Content. Chlorophyll Fluorescence and Photosynthetic Activity of Spruce Clones under Normal and Limited Mineral Nutrition//Photosynthetica. Vol. 27. iss. 3. Pp. 385-400.
- Kowalczyk J. Combining Production of Improved Seeds with Genetic Testing in Seedling Seed Orchards 11 Seed Orchard: Proceedings from a Conference at Umeä. September 26-28. 2007 / ed. by D. Lindgren. Umeä: SLU. 2008. Pp. 126-134.
- Kurteva M, Gateva R. Influence of Industrial and Transport Pollution on the Plastid Pigments of Pinus sylvestris. Picea abies Karst, and Thuja orientalis L. // Scientific Papers of Jubilee Scientific Conference with International Participation “70th Anniversary of the Forest Research Institute”. Sofia. Bulgaria. October 6-7. 1998. Sofia: Forest Research Institute. 1998. Vol. 1. Pp. 146-151.
- Kvicala M, Lackova /-.'., Urbancovd L. Photosynthetic Active Pigments Changes in Norway Spruce (Picea abies) under the Different Acclimation Irradiation and Elevated CO2 Content // ISRN Environmental Chemistry. Vol. 2014. article ID 572576. 4 p. DOI: 10.1155/2014/572576
- Lichtenthaler H.K. Biosynthesis and Accumulation of Isoprenoid Carotenoids and Chlorophylls and Emission of Isoprene by Leaf Chloroplasts I I Bulletin of the Georgian National Academy of Sciences. Vol. 3. no. 3. Pp. 81-94.
- Lichtenthaler H.K, Buschmann C. Chlorophylls and Carotenoids: Measurement and Characterization by UV-VIS Spectroscopy I I Current Protocols in Food Analytical Chemistry. Vol. UNIT F4.3. Pp. F4.3.1-F4.3.8. DOI: 10.1002/0471142913.faf0403s01
- Lichtenthaler H.K, Wellburn A.R. Determinations of Total Carotenoids and Chlorophylls a and b of Leaf Extracts in Different Solvents I I Biochemical Society Transactions. Vol. 11(5). Pp. 591-592. DOI: 10.1042/bst0110591
- Lidholm J., Gustafsson P.A. Functional Promoter Shift of a Chloroplast Gene: A Transcriptional Fusion between a Novel psbA Gene Copy and the //v/MUUU) Gene in Pinus contorta // The Plant Journal. Vol. 2. iss. 6. Pp. 875-886. DOI: 10.1046/j.1365- 313X.1992.tO 1-4-00999.x
- Major J.E., Barsi D.C., Mosseler A., CampbellM. Genetic Variation and Control of Chloroplast Pigment Concentrations in Picea rubens. Picea mariana and Their Hybrids. I. Ambient and Elevated [CO2] Environments // Tree Physiology. Vol. 27(3). Pp. 353-364. DOI: 10.1093/treephys/27.3.353
- Mair as JA. A Review of the Seed Orchard Programme in Poland // Seed Orchard: Proceedings from a Conference at Umeä. September 26-28. 2007 / ed. by D. Lindgren. Umeä: SLU. 2008. Pp. 165-176.
- Niinemets Ü. Acclimation to Low Irradiance in Picea abies: Influences of Past and Present Light Climate on Foliage Structure and Function // Tree Physiology. Vol. 17. iss. 11. Pp. 723-732. DOI: 10.1093/treephys/17.11.723
- Niinemets Ü. Stomatai Conductance Alone Does not Explain the Decline in Foliar Photosynthetic Rates with Increasing Tree Age and Size in Picea abies and Pinus sylvestris // Tree Physiology. Vol. 22. iss. 8. Pp. 515-535. DOI: 10.1093/treephys/22.8.515
- Peguero-Pina J.J., Morales F., Gil-Pelegrin E. Frost Damage in Pinus sylvestris Stems Assessed by Chlorophyll Fluorescence in Cortical Bark Chlorenchyma // Annals of Forest Science. 2008. Vol. 65(8). Pp. 813pl-813p6. DOI: 10.1051/forest:2008068
- Porcar-Castell A., Juurola E., Ensminger I., Berninger F., Hari P., Nikinmaa E. Seasonal Acclimation of Photosystem II in Pinus sylvestris. Using the Rate Constants of Sustained Thermal Energy Dissipation and Photochemistry to Study the Effect of the Light Environment // Tree Physiology. 2008. Vol. 28. iss. 10. Pp. 1483-1491. DOI: 10.1093/treephys/28.10.1483
- Porcar-Castell A., Juurola E., Nikinmaa E., Berninger F., Ensminger I., Hari P. Seasonal Acclimation of Photo system II in Pinus sylvestris. Estimating the Rate Constants of Sustained Thermal Energy Dissipation and Photochemistry // Tree Physiology. 2008. Vol. 28. iss. 10. Pp. 1475-1482. DOI: 10.1093/treephys/28.10.1475
- Rosenthal S.I., Camm E.L. Photosynthetic Decline and Pigment Loss during Autumn Foliar Senescence in Western Larch (Larix occidentalis) // Tree Physiology. Vol. 17. iss. 12. Pp. 767-775. DOI: 10.1093/treephys/17.12.767
- Skuodiene L. Quantitative Changes in Aminoacid Proline and Chlorophyll in the Needles of Picea ahies (L.) during Stress and Adaptation 11 Biologija. 2001. No. 2. Pp. 54-56.
- Wellburn A.R. The Spectral Determination of Chlorophylls a and b. as Well as Total Carotenoids. Using Various Solvents with Spectrophotometers of Different Resolution // Journal of Plant Physiology. Vol. 144. iss. 3. Pp. 307-313. DOI: 10.1016/S0176- 1617(11)81192-2
Похожие статьи
В.П. Бессчетное
Н.Н. Бессчетнова
А.О. Есичев