Эта статья опубликована под лицензией Creative Commons и не автором статьи. Поэтому если вы найдете какие-либо неточности, вы можете исправить их, обновив статью.

Содержание крахмала в тканях побегов разных видов ели (picea a. Dietr.) В условиях интродукции



Н.Н. Бессчетнова
В.П. Бессчетнов
А.В. Кулькова
И.В. Мишукова
Опубликована Март 19, 2017
Последнее обновление статьи Сен. 18, 2022
Эта статья опубликована под лицензией
")



Аннотация
Интродукция является одним из наиболее эффективных методов пополнения местной дендрофлоры ценными видами растений. Для диагностики успешности интродукции растений необходимо выяснить природу их адаптации в новых условиях произрастания. Весьма значимыми для оценки акклиматизации растений являются состав и соотношение запасных веществ и метаболитов в клетках тканей. Из запасных питательных веществ наиболее важен для развития древесных растений крахмал. Повышение температуры значительно влияет на содержание этого питательного вещества. Увеличение содержания крахмала в клетках тканей связано с выходом растений из состояния покоя и подготовкой к вегетационному периоду. Исследована биологическая специфичность 13 видов ели. интродуцированных в Нижегородскую область. Установлена их неоднородность по содержанию крахмала в клетках тканей годичных побегов, что может иметь адаптационное значение. Содержание крахмала в клетках тканей годичных побегов выявляли цветной реакцией с раствором Люголя. Срезы после окрашивания соответствующими реактивами и фиксации анализировали с помощью микроскопа «Микмед-2». Оценку содержания запасных веществ на поперечном срезе побега давали по каждой учетной зоне отдельно и в сумме баллов по всем учетным зонам. Статистический и дисперсионный анализы выполняли по общепринятым методикам. По каждому ботаническому виду, включенному в схему анализа, вычисляли средние значения. Соотношение между видами рода ель по содержанию крахмала в разных тканях было неодинаковым. Различия между интродуцентами проявились на выровненном фоне экологических условий. Дисперсионный анализ подтвердил существенность различий, обнаруженных между исследованными видами ели. в содержании крахмала.
Ключевые слова
Ткани побегов, адаптация, резистентность, ель. интродукция, крахмал
Введение
Интродукция как одна из традиционных в лесном хозяйстве сфер практической деятельности сохраняет свою актуальность и на современном этапе. Большой опыт в направлении активного привлечения многочисленных видов ели в состав различных по целевому назначению и конструкции искусственных насаждений накоплен в Среднем Поволжье, в частности в Нижегородской области [8, 9]. Здесь проходит граница естественных ареалов елей европейской и сибирской, сосредоточены значительные площади естественных еловых насаждений, создано большое количество лесных культур, защитных насаждений и озеленительных посадок этой породы. Широкие возможности применения результатов интродукции в лесокультурном производстве Нижегородской области во многом обусловлены тем, что на ее территории представлена значительная часть лесотипологического разнообразия России, поскольку здесь можно встретить основные природные зоны. В такой ситуации ресурсы адаптации видов неодинаковы, что при организации их изучения выдвигает в число задач оценку соответствия их биологии существующим природным условиям. Привлечение для этих целей методов гистохимических исследований принято достаточно широко [1, 10]. Признаки физиологического состояния растений (морозостойкость, сроки выхода из состояния покоя, скорость роста, сроки макро- и микроспорогенеза и др.) определяют приспособленность организма к среде и его выживаемость [4, 5, 7, 19, 21]. Наличие крахмала в тканях побегов связывают с устойчивостью растений к неблагоприятным зимним условиям. Исследования в территориальном плане по этому направлению проводятся широко и достаточно активно [13-20, 22-25].
С учетом вышеизложенных фактов была определена цель исследований - выявление факта наличия и оценка масштаба межвидовой изменчивости содержания крахмала в тканях побегов различных видов, входящих в таксономическую систему рода ель, для установления видов и форм, наиболее приспособленных к существующему экологическому фону.
Объекты и методы исследования
Объектом исследования служили посадки 13 видов и форм ели, представленных в интродукционной коллекции ботанического сада Нижегородского государственного университета им. Н.П. Лобачевского. Предметом исследования являлась изменчивость содержания крахмала в тканях побегов интродуцированных и аборигенных видов рода ель. Методологической основой работы выступал принцип единственного логического различия, согласно которому в сравнениях участвовали одновозрастные растения, в пределах опытного участка обеспечивалась элиминация дифференцирующего влияния факторов среды, способных вызвать возникновения флуктуаций в проявлениях анализируемых показателей.
Первичная лесоводственная информация была получена в ходе реализации полевых стационарных и лабораторных методов исследования с привлечением общепризнанных методических схем организации работ и построения выборок. В составе обследованного пицетума представлены 13 видов и форм рода ель, различающиеся своими ареалами. Их размещение в границах участка рендоми- зированно. В рамках проводимого исследования им были присвоены следующие условные обозначения: вид 1 - ель Глена (Picea glehnii (F. Schmidt) Mast.); вид 2 - ель канадская (Picea glauca (Moench) Voss); вид 3 - ель шероховатая (Picea asperata Masters); вид 4 - ель черная (Picea mariana Mill., Britton, Stems & Poggenburg); вид 5 - ель колючая, форма серебристая (Picea pungens Engelm., f. argentea); вид 6 - ель колючая, форма голубая (Picea pungens Engelm., f. glauca); вид 7 - ель сербская (Picea omorika (Pancic) Purk.); вид 8 - ель обыкновенная (Picea abies (L.) H. Karst.); вид 9 - ель Зигельмана (Picea engelmannii Parry ex Engelm.); вид 10 - ель колючая (Picea pungens Engelm.); вид 11 - ель аянская (Picea jezoensis (Siebold & Zucc.) Carriere); вид 12 - ель корейская (Picea koraiensis Nakai); вид 13 - ель сибирская (Picea obovata Ledeb.).
В рабочую группу включены только одновозрастные деревья, имеющие одинаковую площадь питания и прочие равные условия произрастания. Количество учетных растений в пределах вида составляло от 1 до 4 в соответствии с их онтогенетической однотипностью и сохранностью на коллекционном участке. С каждого учетного растения заготовлено по 5 нормально развитых однолетних побегов, равномерно размещенных на периферии среднего яруса хорошо освещенного участка кроны. Всего протестировано 145 образцов, заготовленных в марте 2016 г.
Первичной единицей выборки в гистохимическом опыте была принята разовая фиксация количества крахмала в учетной ткани побега на временных препаратах поперечных срезов из средней части годичного прироста. Срезы после окрашивания и фиксации анализировали с помощью микроскопа «Микмед-2». Для анализа и количественной оценки были выделены следующие учетные зоны [3, 7]: сердцевина, перимедуллярная зона ксилемы, сердцевинные лучи, ранняя ксилема, поздняя ксилема, смоляные ходы ранней ксилемы, смоляные ходы поздней ксилемы, смоляные ходы флоэмы, прикамби- альная зона флоэмы, прифеллогенная зона флоэмы, средняя зона флоэмы, корковая зона (зона паренхимных клеток, лежащая на периферии побега между феллогеном и внешней границей побега). Крахмал выявляли цветной реакцией на раствор Люголя [10, 12] и оценивали его содержание в условных баллах по предложенной нами [3, 7] шкале:
0 - крахмал отсутствует полностью, т. е. не удается обнаружить ни одного крахмального зерна ни в одной клетке учетной зоны ткани;
1 - крахмал в ограниченном количестве присутствует в единичных (до 10 %) клетках учитываемой ткани;
2 - крахмал присутствует в меньшем, чем половина, числе клеток, окрашенные крахмальные зерна удается зафиксировать примерно у 20... 30 % клеток;
3 - крахмал фиксируется примерно в половине (40... 60 %) всех учитываемых клеток ткани;
4 - крахмал отмечается в большей части (70... 90 %) клеток;
5 - крахмал в большом количестве присутствует во всех клетках (100 %) учитываемой ткани.
Оценку давали по каждой учетной ткани отдельно и в сумме баллов по всем учетным зонам. По каждому ботаническому виду, включенному в схему анализа, вычисляли средние значения. В качестве контроля визирования использовали неокрашенные срезы, не подвергавшиеся воздействию тестирующих реагентов [2, 3, 7]. С ними сравнивали срезы, обработанные соответствующими реактивами. Окраска их клеток указывает на отсутствие запасных веществ. Гистохимический анализ выполняли в лаборатории кафедры лесных культур Нижегородской государственной сельскохозяйственной академии, статистический и дисперсионный анализы - по общепринятым схемам, а также с применением алгоритмов, разработанных авторами [6, 7]. Для оценки уровней изменчивости анализируемых показателей использовали шкалу С.А. Мамаева [11].
Результаты исследования и их обсуждение
В ходе проведения опытов установлены хорошо заметные различия между анализируемыми видами ели по содержанию крахмала в клетках тканей 1-летних побегов (табл. 1-3).
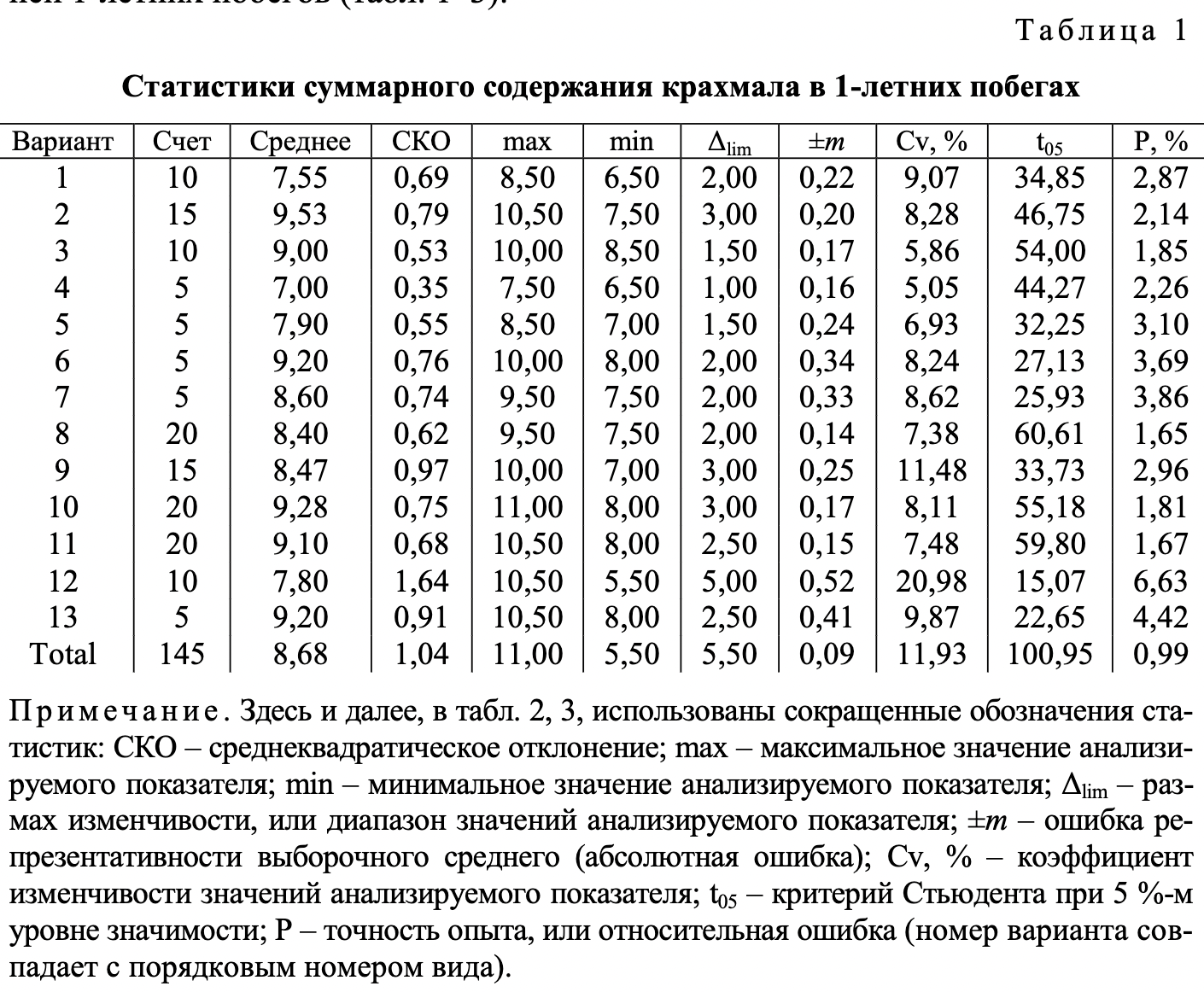
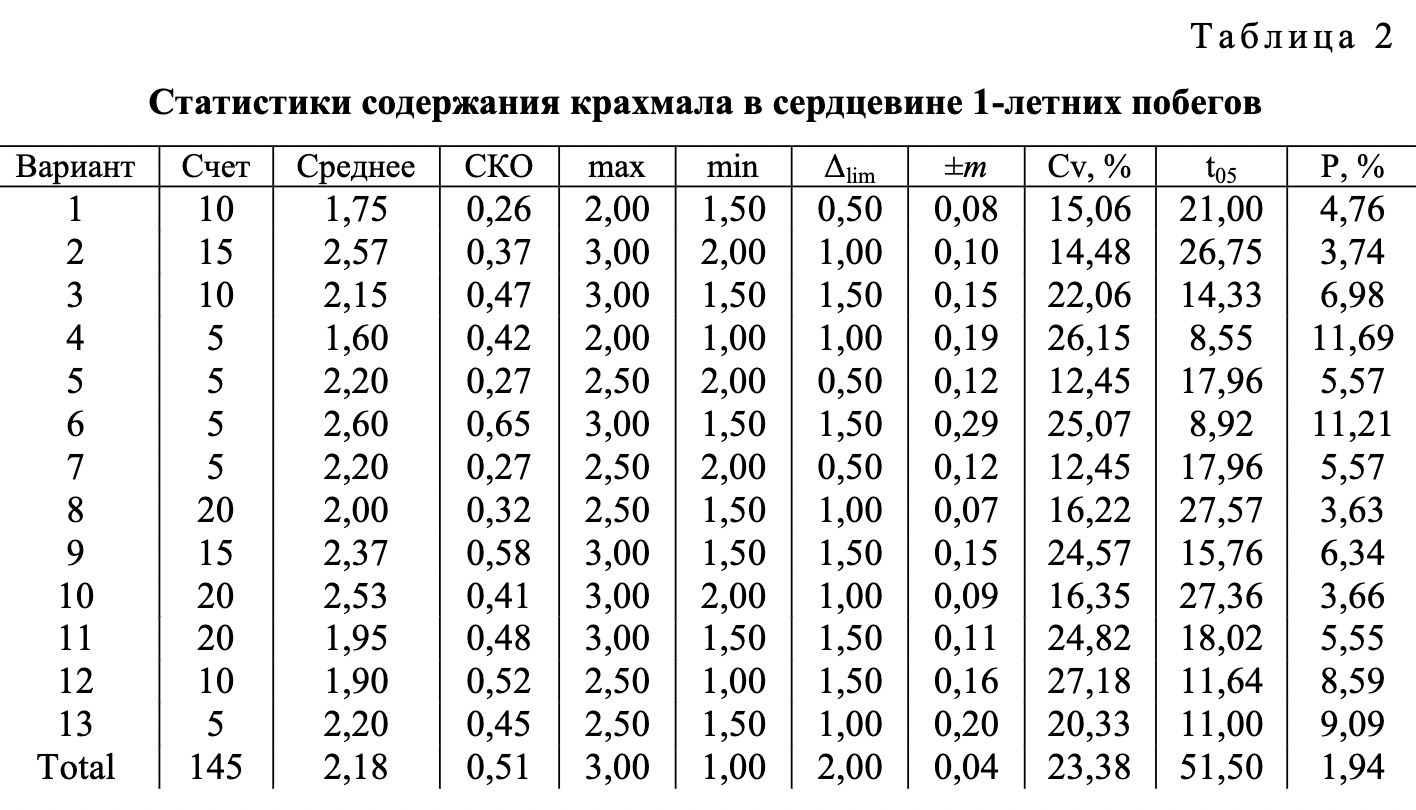
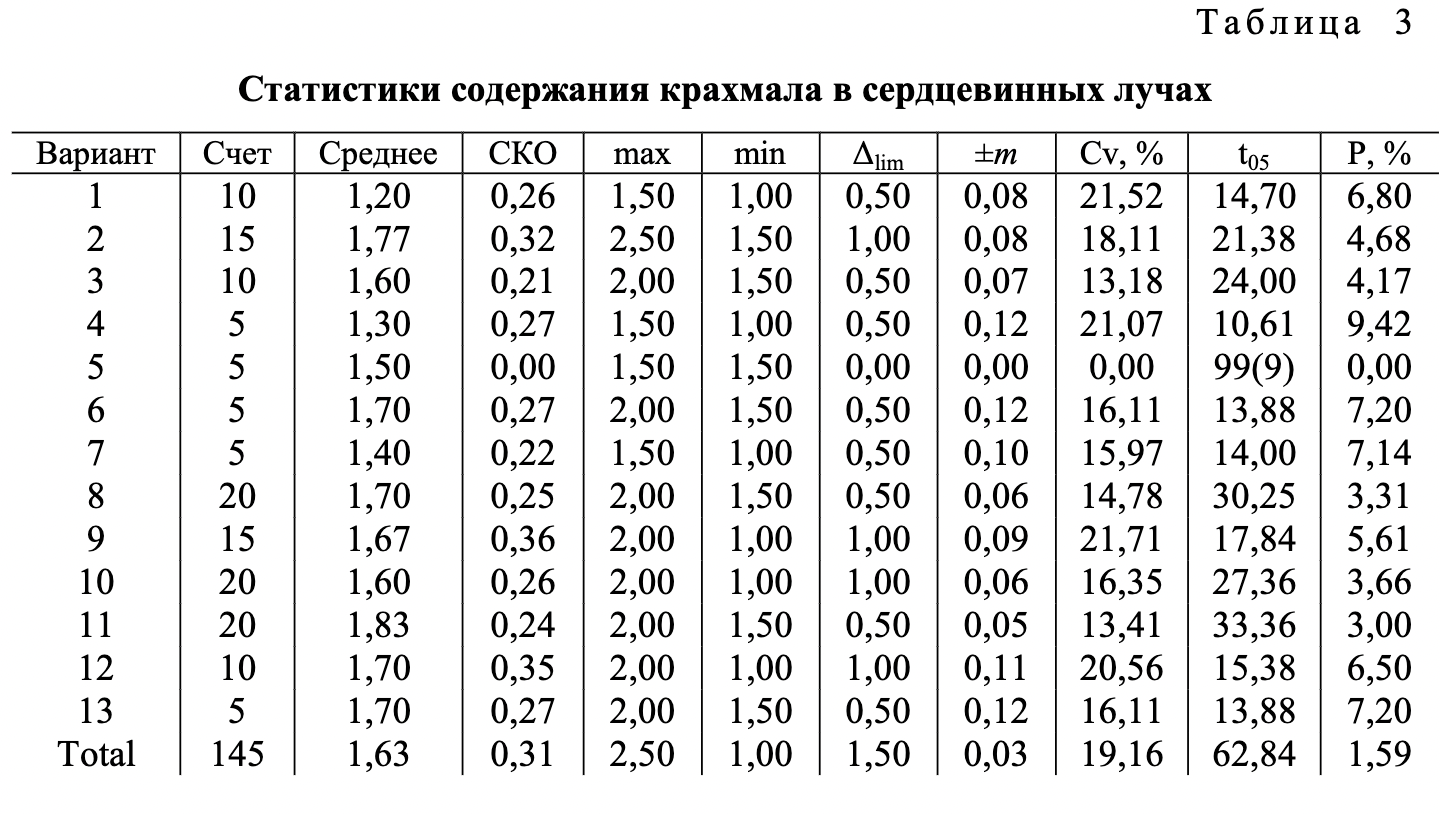
Полученный в ходе статистической обработки материал достоверен и вполне надежен. Опытные t-критерии Стьюдента намного превышают критическое значение, равное 1,96 при 5 %-м уровне значимости. Показатели точности опыта (относительная ошибка Р) в анализе суммарного количества крахмала не преодолевают допустимый уровень в 5 %. Вместе с тем в анализе содержания крахмала по отдельным тканям в ряде случаев они оказываются выше указанного предела, что связано с ограниченным числом определений при существующем уровне дисперсии признаков.
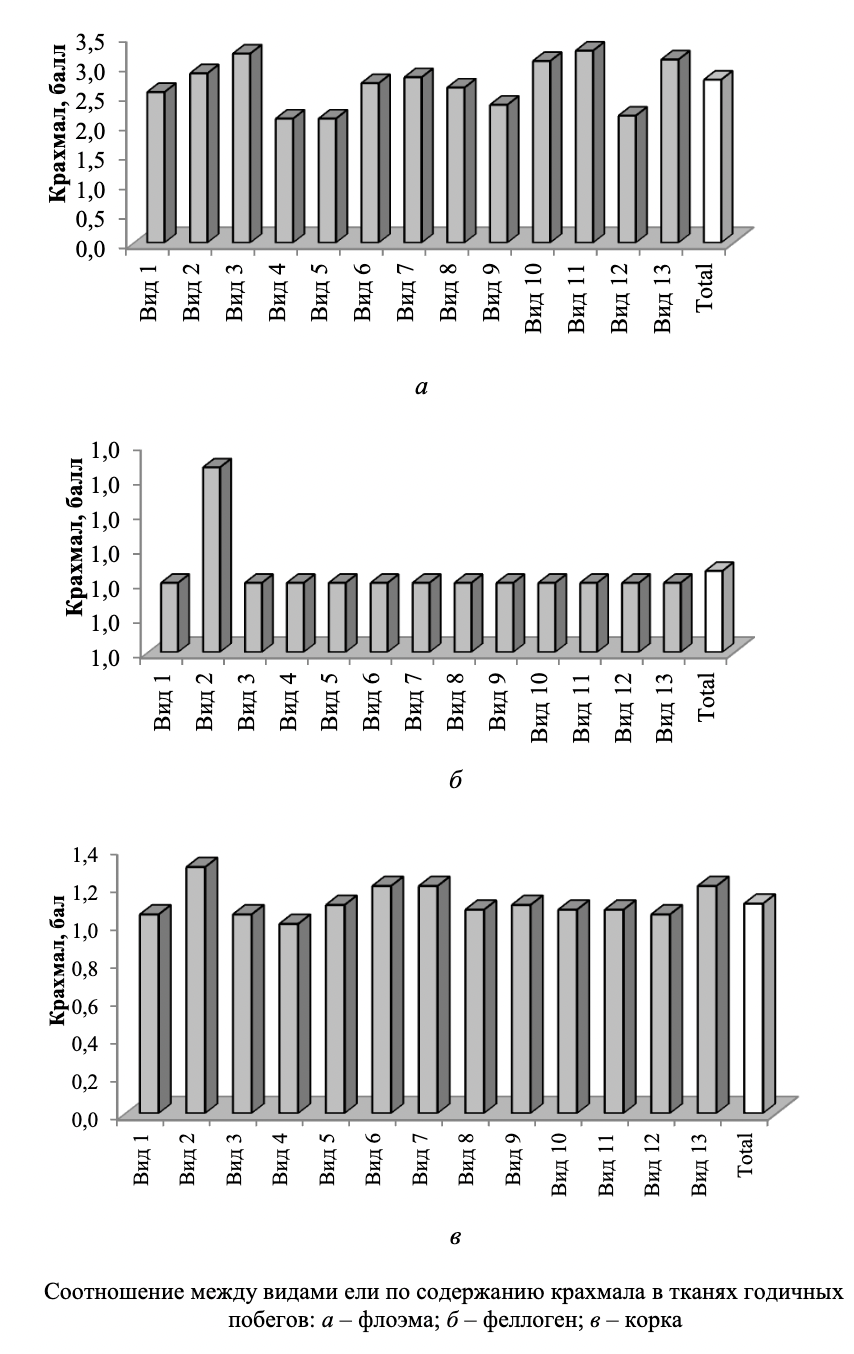
Оценки изменчивости анализируемых показателей по шкале С.А. Мамаева в обобщенном массиве данных соответствуют низкому и среднему уровню: суммарное содержание крахмала Сѵ = 11,93 % (низкий); содержание крахмала в сердцевине Сѵ - 23,38 % (средний); содержание крахмала в сердцевинных лучах Сѵ — 19,16 % (средний).
В анализируемом составе видов ели средние значения изучаемых характеристик были достаточно стабильны. В частности, по суммарному содержанию крахмала наибольшее среднее превосходило наименьшее в 1,36 раза; по содержанию крахмала в сердцевине - в 1,62 раза; по содержанию крахмала в сердцевинных лучах - в 1,52 раза. Соотношение абсолютных значений указанных показателей было несколько выше и соответственно составляло: в 2,0; 3,0 и 2,5 раза.
Наглядное представление о соотношении между рассматриваемыми видами ели по показателю наличия крахмала в разных тканях 1-летних побегов дают диаграммы (см. рисунок). Зафиксированные между исследуемыми видами различия в содержании крахмала в тканях побегов проявились на фоне выровненных условий произрастания, что позволяет признать причиной их возникновения межвидовую изменчивость.
Достоверность указанных различий была подтверждена результатами однофакторного дисперсионного анализа (табл. 4).
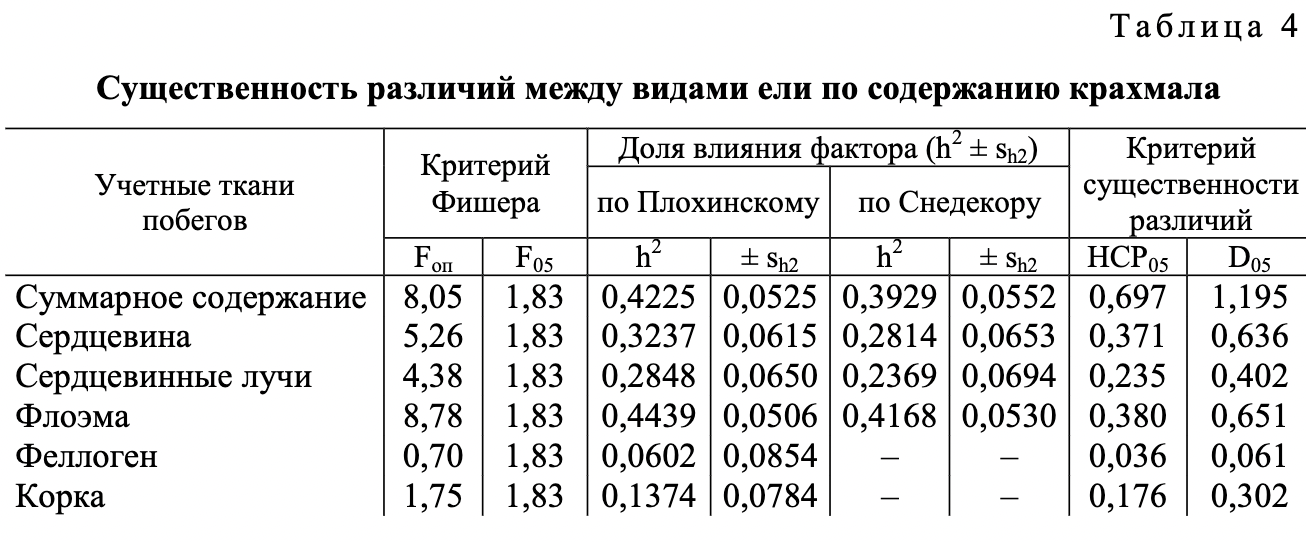
Сравниваемые образцы существенно различались между собой по содержанию крахмала в большинстве изученных тканей побегов. В случаях с подтвержденной существенностью различий опытные В-критерии Фишера заметно больше соответствующих табличных значений при 5 %-м уровне значимости.
Не установлено наличие существенных различий по содержанию крахмала в феллогене и корковой зоне. Здесь значения критерия Фишера оказались меньше критических значений. Влияние собственно различий между видами в вариантах опыта с подтвержденной существенностью различий достоверно и значительно. Суммарное содержание крахмала в тканях побегов в расчетах
по алгоритму Плохинского составило (42,25±5,25) %. А эффект по ряду тканей был несколько ниже: (32,37±6,15) % (сердцевина) и (28,48±6,50) % (сердцевинные лучи). При этом оценки по флоэме оказались выше: (44,39±5,06) %. Во всех вариантах опыта замечено преобладающее влияние факторов среды, которое формирует остаточную дисперсию. На его долю приходится от 55,61 % (флоэма) до 93,98 % (феллоген) общего эффекта.
Оценки, полученные с помощью алгоритма Снедскора, оказались близкими по значениям. Критерии существенности различий (НСР05 и D,)S) позволяют установить, между какими именно видами различия по анализируемым показателям (см. табл. 1-3) могут быть причислены к категории существенных. В частности, по суммарному содержанию крахмала ель черная существенно отличалась от остальных 9 видов, участвующих в испытании. Ели Глена и корейская имели по 6 существенных различий с другими видами. Оставшиеся виды более сходны между собой по данному показателю. Так, ели сербская, европейская и Зигельмана проявили существенные различия только в одном случае их парных сравнений с другими.
Заключение
Полученный материал позволил выделить группы наиболее сходных ранней весной по физиологическому состоянию видов елей. Данный показатель является индикатором завершения сроков заготовки черенков елей для размножения. Известно, что одревесневшие черенки демонстрируют более высокие темпы укоренения при их заготовке в состоянии покоя. Очевидно, что виды, обладающие в середине марта достаточно высоким запасом крахмала, характеризуются более ранними сроками начала вегетации и, следовательно, более ранними сроками завершения заготовки черенков. К таким отнесены следующие виды ели: канадская (Picea glauca (Moench) Voss); шероховатая (Picea asperata Masters); колючая, форма голубая (Picea pungens Engelm., / glauca); колючая (Picea pungens Engelm.); аянская (Picea jezoensis (Siebold & Zucc.) Carriere); сибирская (Picea obovata Ledeb.).
СПИСОК ЛИТЕРАТУРЫ
- Барская Е.П. Изменения хлоропластов и вызревание побегов в связи с морозоустойчивостью древесных растений. М.: Наука. 1967. 223 с.
- Бессчеттюва НН. Сравнительная оценка плюсовых деревьев сосны обыкновенной по содержанию крахмала в побегах // Вести. МарГТУ. Серия: Лес. Экология. Природопользование. 2010. № 2(9). С. 49-56.
- Бессчептова Н.Н. Сосна обыкновенная (Pinus sylvestris). Селекционный потенциал плюсовых деревьев. Саарбрюккен: Lap Lambert Academic Publishing. 2011. 402 c.
- Бессчептова Н.Н. Содержание жиров в клетках побегов плюсовых деревьев сосны обыкновенной // Леей. жури. 2012. № 4. С. 48-55. (Изв. высш. учеб, заведений).
- Бессчетное В.П., Бессчетнова Н.Н. Образование и лигнификация ксилемы плюсовых деревьев сосны обыкновенной // Леси. жури. 2013. № 2. С. 45-52. (Изв. высш. учеб, заведений).
- Бессчетное В.П., Бессчетнова Н.Н., Храмова О.Ю., Орнатский А.Н., Борелов Н.П. Дисперсионный анализ многоуровневых иерархических комплексов: метод, указания для студ. и асп. Н. Новгород: НГСХА. 2012. 33 с.
- Бессчетнова Н.Н. Сосна обыкновенная (Pinus sylvestris). Эффективность отбора плюсовых деревьев: моногр. Н. Новгород: Нижегород. ГСХА. 2016. 464 с.
- Воробьев Р.А., Тебенькова Д.Н. Развитие вегетативных и генеративных органов представителей рода ель (Picea). интродуцированных в Нижегородской области//Вести. МГУЛ-Леей, вести. 2013. № 7. С. 97-105.
- Воробьев Р.А., Тебенькова Д.Н. Содержание основных пигментов в однолетней и двухлетней хвое интродуцентов рода ель (Picea) в условиях подзоны южной тайги (на примере Нижегородской области) // Лесоведение. 2013. №3. С. 8-15.
- Бродзинский А.М., Рродзинский Д.М. Краткий справочник по физиологии растений. Киев: Наук, думка. 1964. 288 с.
- Мамаев С.А. О проблемах и методах внутривидовой систематики растений.
- Амплитуда изменчивости // Закономерности формообразования и дифференциации вида у древесных растений: тр. Ин-та экологии растений и животных Урал. фил. АН СССР. Свердловск. 1969. Вып. 64. С. 3-38.
- Прозина М.Н. Ботаническая микротехника: учеб, пособие. М.: Высш. шк. 1960. 205 с.
- Ander sone U., levinsh G. Changes of Morphogenic Competence in Mature Pinus svlvestris Buds in vitro. Ann. Bot. 2002. Vol. 90. no. 2. pp. 293-298. DOI: 10.1093/aob/mcfl76
- Bergstrom B. Chemical and Structural Changes during Heartwood Formation in Pinus sylvestris // Forestry. Vol. 76. no. 1. Pp. 45-53. DOI: 10.1093/forestry/76.1.45.
- Brahim B.M., Loustau D., Gaudi Here J.P., Saur E. Effects of Phosphate Deficiency on Photosynthesis and Accumulation of Starch and Soluble Sugars in 1-Year-Old Seedlings of Maritime Pine (Pinus pinaster Ait) // Ann. Sei. For. 1996. 53. no. 4. Pp. 801-810. DOI: http://dx.doi.Org/10.1051/forest:19960401
- Ericsson A., Lindgren A., Mattsson A. Effects of Cold-Storage and Planting Date on Subsequent Growth. Starch and Nitrogen Content in Scots Pine (Pinus svlvestris) and Norway Spruce (Picea abies) Seedlings I I Studia Forestalia Suecia. No. 165. Pp. 1-17.
- Jimenez M.D., Pardos M., Puertolas J., Kleczkowski L.A., Pardos J.A. Deep Shade Alters the Acclimation Response to Moderate Water Stress in Quercus suber // Forestry. 2009. Vol. 82. no. 3. Pp. 285-298. DOI: 10.1093/forestry/cpp008
- Kakei M„ Clifford P.E. Effects of Lime Application and Test Branch Position on 14C-Photosynthate Partitioning in 3-Year-Old Sitka Spruce (Picea sitchensis) Plants Growing in Pots Containing Peat Soils // Forestry. Vol. 72. no. 3. Pp. 223-235. DOI: 10.1093/forestry/72.3.223
- Ludlow A.R. Some Factors Influencing the Increment of Forests // Forestry. Vol. 70. no. 4. Pp. 381-388. D01:10.1093/forestry/70.4.381
- Mencuccini M, Hölttä T. The Significance of Phloem Transport for the Speed with which Canopy Photosynthesis and Belowground Respiration are Linked // New Phytol. Vol. 185. no. 1. Pp. 189-203. DOI: 10.1111/j.l469-8137.2009.03050.x
- Mencuccini M., Hölttä T., Sevan to S., Nikinmaa E. Concurrent Measurements of Change in the Bark and Xylem Diameters of Trees Reveal a Phloem-Generated Turgor Signal//New Phytol. Vol. 198.no. 4.Pp. 1143-1154. DOI: 10. Ill 1/nph. 12224
- Pardos AL, RoyoA., Gil L., Pardos JA. Effect of Nursery Location and Outplant- ing Date on Field Performance of Pimis halepensis and Quercus ilex Seedlings // Forestry. Vol. 76. no. 1. Pp. 67-81. DOI: 10.1093/forestiy/76T.67
- Rodriguez-Calcerrada J, Pardos JA., Aranda 1. Contrasting Responses Facing Peak Drought in Seedlings of Two Co-Occurring Oak Species // Forestry. Vol. 83, no. 4. Pp. 369-378. D01:10.1093/forestry/cpq019
- Schaberg P.G., Snyder M.C., Shane J.B., Donnelly J.R. Seasonal Patterns of Carbohydrate Reserves in Red Spruce Seedlings I I Tree Physiology. Vol. 20. no. 8. Pp. 549-555.
- Woodruff D.R., Meinzer F.C. Water Stress. Shoot Growth and Storage of Non- structural Carbohydrates Along a Tree Height Gradient in a Tall Conifer // Plant. Cell & Environment. 2011. Vol. 34. no. 11. Pp. 1920-1930. DOI: 10.1111/j.l365- 3040.2011.02388.x
Похожие статьи
В.П. Бессчетное
Н.Н. Бессчетнова
А.О. Есичев
Виктория Викторовна Руженкова
Герасимова Ольга Владимировна
Белоусова Ольга Викторовна
Белоусов Евгений Александрович
Яницкий Олег Николаевич
Кубасов Р.В.