This article is published under a Creative Commons license and not by the author of the article. So if you find any inaccuracies, you can correct them by updating the article.

Species distribution and depth habitat of recent planktic foraminifera in Fram Strait, Arctic Ocean



Theodora Pados
Robert F. Spielhagen
Published: May 27, 2014
Latest article update: Aug. 21, 2023
This article is published under the license




Abstract
To describe the horizontal and vertical distribution of recent planktic foraminifera in Fram Strait (Arctic), plankton samples were collected in the early summer of 2011 using a MultiNet sampler (>63 µm) at 10 stations along a west–east transect at 78°50′N. Five depth intervals were sampled from the sea surface down to 500 m. Additionally, sediment surface samples from the same locations were analysed. The ratio between absolute abundances of planktic foraminifera in the open ocean, at the ice margin and in the ice-covered ocean was found to be approximately 2:4:1. The assemblage was dominated by the polar Neogloboquadrina pachyderma (sin.) and the subpolar Turborotalita quinqueloba, which accounted for 76 and 15% of all tests in the warm, saline Atlantic waters and 90 and 5% in the cold and fresh Polar waters, respectively. Both species had maximum absolute abundances between 0 and 100 m water depth, however, they apparently lived shallower under the ice cover than under ice-free conditions. This indicates that the depth habitat of planktic foraminifera in the study area is predominantly controlled by food availability and not by temperature. The distribution pattern obtained by plankton tows was clearly reflected on the sediment surface and we conclude that the assemblage on the sediment surface can be used as an indicator for modern planktic foraminiferal fauna.
Keywords
Fram Strait, Arctic Ocean, N. pachyderma (sin.), T. quinqueloba, Planktic foraminifera, depth habitat
Planktic foraminifera are protozoa that inhabit the upper part of the water column in the world oceans. As their species distribution in high latitude oceans reflects properties of their habitat (i.e., water temperatures and the position of the summer sea-ice margin), fossil assemblages in sediments are often used to reconstruct palaeoenvironments. However, for a correct interpretation of fossil data, it is important to improve our understanding of the correlation between environmental variability in the ocean and the related distribution of living foraminifera. It is particularly essential to reveal the actual depth habitat of recent specimens, as the water properties reconstructed from their fossil calcite shells must be attributed to correct water depths.
Many studies on recent planktic foraminiferal distributions were conducted in tropical/subtropical areas (e.g., Bé & Hutson 1977; Andrijanic 1988; Naomi et al. 1990; Kuroyanagi & Kawahata 2004), while rather few studies concentrated on (sub-)Arctic regions, mainly on Fram Strait. Hansen & Knudsen (1995) published data from Freemansundet, Svalbard, Carstens & Wefer (1992) presented results on recent foraminifera in the Nansen Basin, and Kohfeld et al. (1996) analysed samples from the Northeast Water Polynya. Close to our work area, studies have been conducted by Carstens et al. (1997) and Volkmann (2000), who showed that planktic foraminiferal species composition in Fram Strait is dominated by polar Neogloboquadrina pachyderma (sinistral) (Ehrenberg 1861) and subpolar Turborotalita quinqueloba (Natland 1938). The abundance ratio of these two species varies with the hydrographical regime but they both occur in the upper 500 m of the water column, with highest abundances at the sea-ice margin. In this paper, we present new data on the vertical and horizontal distribution of planktic foraminifera species, collected during the Polarstern ARK-XXVI/1 cruise, in 2011, at 10 stations along a transect at 78°50′N across Fram Strait (Fig. 1). The samples were taken using a MultiNet sampler with nets of 63 µm mesh size at five depth intervals. Additionally, the sediment surface at the same locations was sampled. In this area, the production maximum of planktic foraminifera usually occurs in mid-summer (Kohfeld et al. 1996; Jonkers et al. 2010). Foraminifer assemblages in core-top and down core samples from Fram Strait are therefore thought to represent modern and past summer conditions, respectively. In order to examine whether processes during deposition (e.g., lateral transport, selective dissolution) significantly change the distribution of planktic foraminiferal tests in sediments, our MultiNet sampling results are also compared to the planktic foraminifer assemblages found on the sea floor.
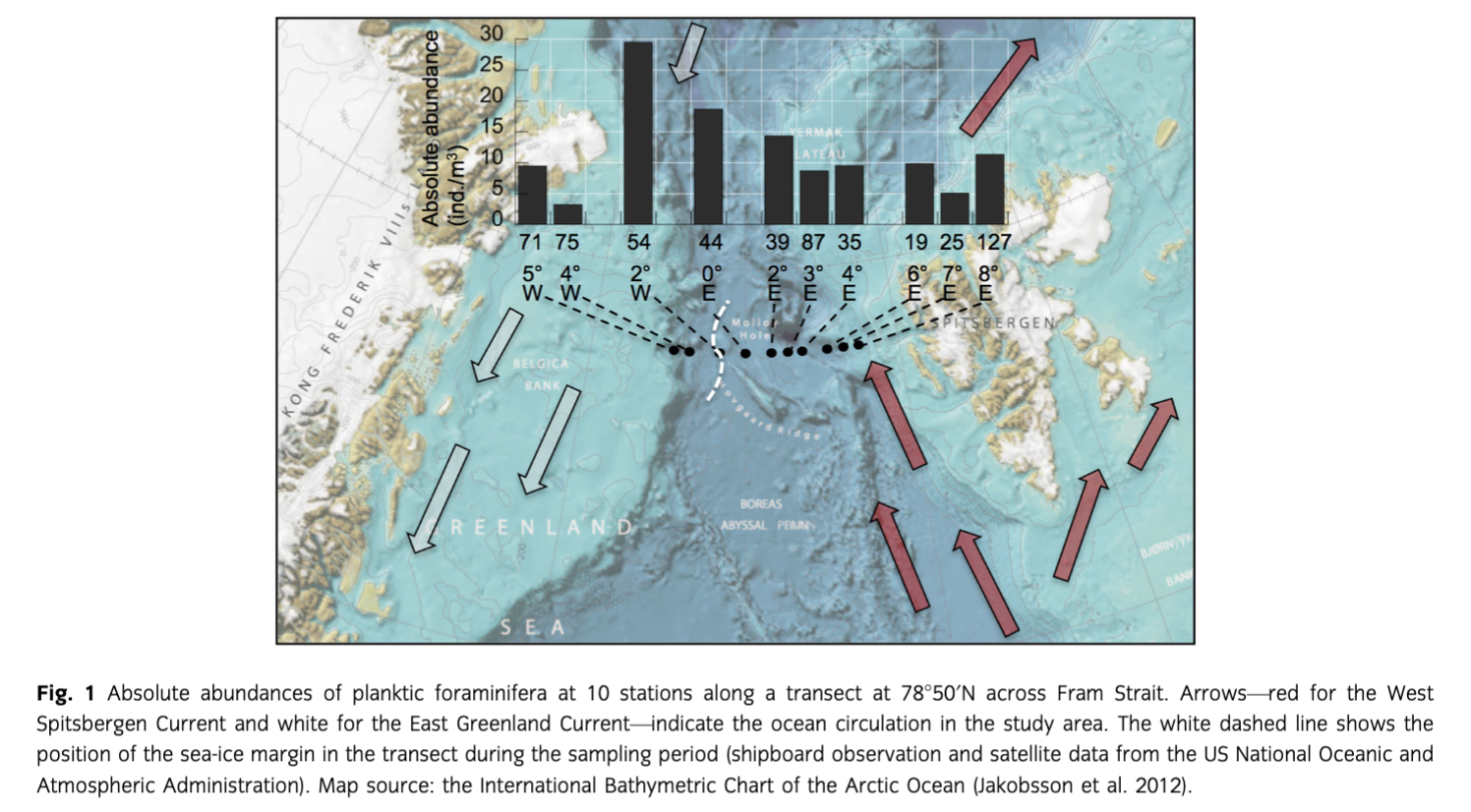
Oceanography
Fram Strait is characterized by high oceanographic variability. Two major surface current systems dominate the area: the West Spitsbergen Current (WSC) in the eastern part that transports water masses northward and the southward flowing East Greenland Current (EGC) in the west (Johannessen 1986). The upper 500 m of the WSC is dominated by Atlantic Water with a thin (<5 m) mixed layer on top in the summer, resulting from ice melting. In winter, the Polar Surface Water takes over its place and forms a thicker wedge to the west (Rudels et al. 1999). The Atlantic Layer is characterized by salinities above 34.5 and temperatures higher than 0.5°C. It ranges between 200 and 500 m. The Polar Mixed Layer (PML) is colder, with a low salinity (<34) and about 50 m thickness. The two layers are isolated by a cold halocline (Jones 2001). The EGC carries cold, low-saline Arctic outflow waters in the upper 200 m. As a result of ice melt, surface waters at the East Greenland continental slope are characterized by extremely low salinities in summer (below 32). The PML here is located above a well-developed halocline (Rudels et al. 2000). Underneath the halocline, warmer and saline waters of Atlantic origin can be found. The sea-ice extent in the strait is controlled by the interaction between Polar and Atlantic waters and is highly variable (Vinje 1977, 2001): usually only the shelf of East Greenland and the northern Fram Strait stay covered by ice in the summer, but in the case of extremely cold winter/spring periods the ice can even intrude into the eastern part of the strait (Dickson et al. 2000).
Oceanographic measurements during ARK-XXVI/1 cruise (Beszczynska-Möller & Wisotzki 2012) provided a detailed profile across Fram Strait at the time of sampling (Fig. 2). The Atlantic Layer in the east was characterized by salinities near 35 and temperatures between 7 and 2°C, while the Polar Layer in the western part of the strait yielded salinities around 33 and a mean temperature of −1.5°C. Here, on the surface Beszczynska-Möller & Wisotzki (2012) found the above-mentioned low-salinity layer with lowest values of 30.6. Underneath the Arctic-derived and mainly ice-covered water mass, submerged warm and salty Atlantic waters can be found. The sea-ice margin in the transect was located at ca. 2°W during the sampling period.

Material and methods
Samples used in this study were obtained during the ARK-XXVI/1 expedition with the research vessel Polarstern in late June/early July 2011 in Fram Strait. Plankton samples were collected by a MultiNet sampler (net opening 0.5 m2; Hydro-Bios, Kiel, Germany) at 10 stations along a transect at 78°50′N across Fram Strait (Fig. 1), (Table 1). The nets of 63 µm mesh size were towed vertically on regular depth intervals (500–300 m, 300–200 m, 200–100 m, 100–50 m, 50–0 m) with a maximum winch speed of 0.3 m/s. During the sampling, the volume of filtered water was measured with a flow metre attached to the MultiNet frame. The flow metre was discovered to be defective and could not be repaired during the cruise. For some MultiNet casts and depth intervals, the instrument gave no or obviously unreliable results. We have therefore decided to omit the flow metre data for this study. The filtered volume was calculated from the mouth opening of the MultiNet and the vertical length of the towed interval for each sample. The plankton samples were sieved with a sieve of 500 µm mesh size, fixed with ethanol and stored at 4°C on board. Before picking the foraminifera by a pipette, all samples were rinsed with distilled water, treated with hydrogen peroxide to remove organic material and preserved in seawater. The picked specimens were rinsed with distilled water, dried on air and ashed in a low temperature vacuum asher for 90 min.
Table 1 List of the stations sampled during the ARKXXVI/1 cruise with the Polarstern in June/July 2011. | |||||
Station no. | Latitude | Longitude | Water depth | Date | Ice cover |
PS78-19 | 78°49.84′N | 6°0.69′E | 2464 m | 25.06.11 | No |
PS78-25 | 78°49.962′N | 7°0.077′E | 1465 m | 26.06.11 | No |
PS78-35 | 78°49.772′N | 3°58.380′E | 2335 m | 28.06.11 | No |
PS78-39 | 78°50.09′N | 1°54.56′E | 2554 m | 28.06.11 | No |
PS78-44 | 78°49.972′N | 0°4.630′E | 2636 m | 29.06.11 | No |
PS78-54 | 78°50.02′N | 2°0.21′W | 2714 m | 01.07.11 | Ice margin |
PS78-71 | 78°49.66′N | 5°20.99′W | 684 m | 04.07.11 | Ice covered |
PS78-75 | 78°49.74′N | 3°55.44′W | 1978 m | 04.07.11 | Ice covered |
PS78-87 | 78°50.44′N | 3°0.19′E | 2454 m | 06.07.11 | No |
PS78-127 | 78°49.84′N | 8°1.33′E | 1019 m | 10.07.11 | No |
Sediment surface samples were obtained from multicorer deployments at the same stations and preserved in ethanol. Samples were freeze-dried, wet-sieved with distilled water through a 63 µm mesh, dried at 40°C and split into several fractions.
All foraminifera from a split of the 100–250 µm fraction of the plankton samples (half or quarter) and from a split of the 100–250 µm fraction of the sediment surface samples (containing >300 individuals) were identified using the taxonomy of Hemleben et al. (1989) and counted under dry conditions. Temperature and salinity of the water column were measured by a conductivity–temperature–depth (CTD) profiler (Beszczynska-Möller & Wisotzki 2012) immediately before the plankton tows. Data of ice coverage were obtained by shipboard observations.
Results
Planktic foraminifera in the plankton samples
The absolute abundances of planktic foraminifera of the 100–250 µm size class in the upper 500 m of the water column show a general increase from ca. 10 individuals per cubic metre (ind./m3) at the eastern stations to 29 ind./m3 at the sea-ice margin (station 54). Further westward we can see a drastic decrease (3 and 9 ind./m3, stations 75 and 71, respectively; Fig. 1).
The species assemblage is dominated by polar Neogloboquadrina pachyderma (sin.) and subpolar Turborotalita quinqueloba, with proportions of around 76 and 15% in the east and 90 and 5% in the west, respectively. Other planktic foraminifera species found in the samples (N. pachyderma [dex.] and Globigerina bulloides) contribute less than 9% to the faunal composition. The relative abundance of N. pachyderma (sin.) varies between 73 and 92% with an increasing trend towards the west, while T. quinqueloba shows a reverse tendency with the highest value of 23% in the eastern part of the strait and the lowest (5%) at the Greenland slope. Figure 3 shows the species composition at the stations in preselected depth intervals. N. pachyderma (sin.) contributes less than 70% to the total planktic foraminiferal fauna at two stations (at 8°E and 6°E), between 0 and 50 m water depth. The highest proportion of this species was found at station 75 (4°W).

The two dominant species, N. pachyderma (sin.) and T. quinqueloba, show similar trends in the depth distribution: both have maximum absolute abundances (ind./m3) between 0 and 100 m water depths (Fig. 4). N. pachyderma (sin.) has highest abundances near the surface between 0 and 50 m at stations 35 and 87 in the middle of the strait (4°E and 3°E, respectively) and under the ice cover at station 75. Highest amounts of T. quinqueloba in the upper 50 m were found at stations 127, 35 and 44 (8°E, 4°E and 0°E, respectively); at the remaining stations, the depth of 50–100 m yielded maximum abundances of this species.
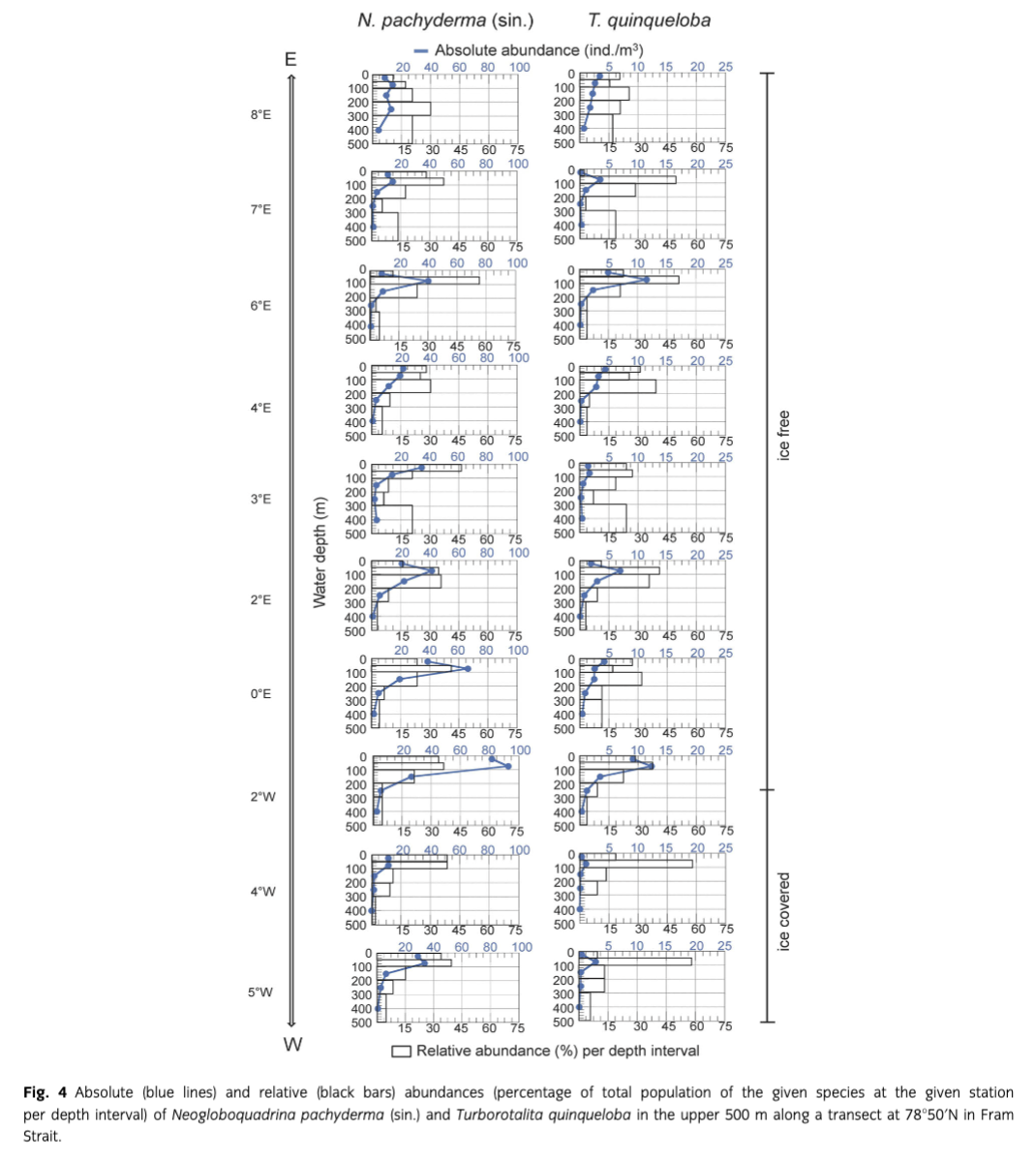
Looking at the relative abundances (percentage of total population of the given species at the given station per depth interval; Fig. 4), both species show a trend to increasing percentages from the surface to the interval 50–100 m, followed by a decrease to 300–500 m. Only at the easternmost station (127), the population maximum of N. pachyderma (sin.) descends below 200 m, while the highest relative abundance of T. quinqueloba at all stations lies above 200 m.
Sediment samples
The samples do not show any evidence of selective carbonate dissolution. Only few broken shells were found, and the two studied species do not show any significant difference in the degree of fragmentation.
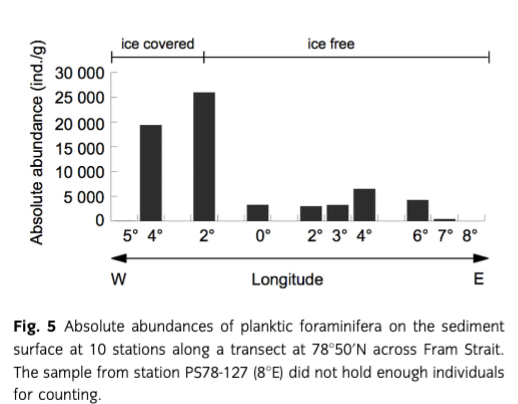
In the absolute abundances of planktic foraminifera, we see a trend similar to that of the plankton samples: the station close to Svalbard (25) has a low value (<400 ind./g sediment), followed by a minor peak at 4°E (ca. 6500 ind./g). After a steady interval (ca. 3000 ind./g), a significant peak can be observed at the sea-ice margin (station 54, ca. 26 000 ind./g), followed by a drastic decrease to ca. 50 ind./g below the sea ice (Fig. 5).
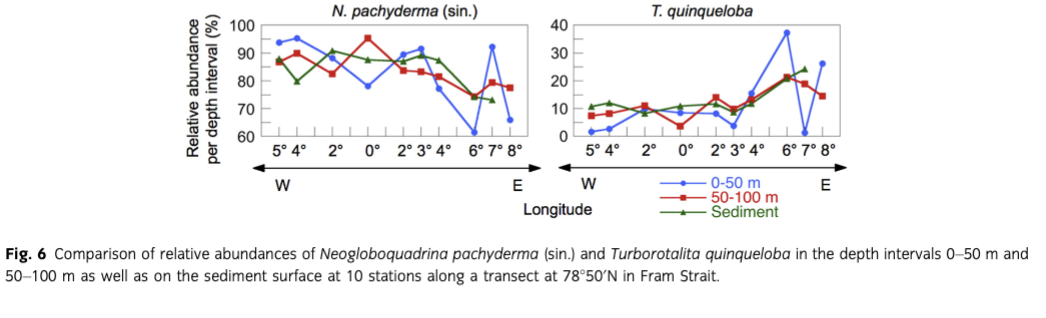
Neogloboquadrina pachyderma (sin.) has highest proportions in the assemblage at the sea-ice margin (91%) and lowest (73%) in the east (7°E, station 25). Between these two stations, a steady increase can be detected. T. quinqueloba shows an exactly reverse trend with abundances between 8 and 24% (Fig. 6). Comparing the relative abundances of N. pachyderma (sin.) and T. quinqueloba on the sediment surface to those counted in the upper 100 m of the water column, the species composition in the depth interval of 50–100 m follows the trend of the core-top samples more precisely than the depth interval between 0 and 50 m mirrors the sediment surface (Fig. 6). All count data can be found in Supplementary Tables S1–S3.
Discussion
Species assemblage
The planktic foraminiferal species composition of the study area resembles the typical foraminiferal fauna previously reported from Arctic/sub-Arctic regions with the clear dominance of Neogloboquadrina pachyderma (sin.) and Turborotalita quinqueloba. A similar species distribution has been already shown for the Arctic Ocean (Bé 1960; Carstens & Wefer 1992) and for Fram Strait (Carstens et al. 1997; Volkmann 2000). Furthermore, despite the warming of Atlantic waters passing through the strait (Spielhagen et al. 2011), the polar species N. pachyderma (sin.) still clearly prevails in our samples, compared to the subpolar T. quinqueloba. Interestingly, at the sampled stations, T. quinqueloba showed relatively low values compared to the results of the above-mentioned studies (Fig. 3). Carstens et al. (1997) observed a range from 15 to 64% for this species, while the proportion of T. quinqueloba in certain samples collected by Volkmann (2000) in warm, Atlantic Water-dominated regimes reached even 93%. In our plankton samples, the relative abundance of T. quinqueloba varied between 5 and 23% and the sediment surface samples showed a very similar trend. These discrepancies point to a variable foraminiferal production found due to different sampling periods. Carstens et al. (1997) and Volkmann (2000) took their samples in August, while our samples were collected in late June/early July and therefore probably reflect an early summer plankton bloom. In the eastern North Atlantic Ocean, Schiebel & Hemleben (2000) found distinctly different planktic foraminiferal faunas in June and in August. In contrast to early summer, the late summer assemblage was dominated by species preferring warmer waters than other species present, just like the subpolar T. quinqueloba in the planktic foraminiferal fauna of Fram Strait. It has been shown for the western North Atlantic that, while the production of N. pachyderma (sin.) has two peaks during the season and one already in early summer, the production of T. quinqueloba reaches its maximum only in early autumn, after the second bloom of N. pachyderma (sin.) (Jonkers et al. 2010). Although sediment trap studies in the Northeast Water Polynya revealed a unimodal pattern of planktic foraminiferal production (Kohfeld et al. 1996), our data indicate that a temporal offset in the production maxima of the two species exists also at these northern latitudes. This could lead to the relatively low numbers of T. quinqueloba in our samples, collected in June/July compared to the studies conducted in August.
Horizontal distribution
The variable species composition along the transect reflects the complex hydrography of Fram Strait. In the east, the WSC carries warm, saline Atlantic Water northwards. In this area, the subpolar T. quinqueloba had higher abundances and the polar N. pachyderma (sin.) had lower abundances than in the western part where the upper 200 m of the EGC consist of cold, low-saline Arctic outflow waters. Here, N. pachyderma (sin.) had a proportion of around 90%. This indicates that the horizontal distribution of the two species is primarily controlled by the distribution of different water masses.
The ratio between absolute abundances of planktic foraminifera in the open ocean, at the ice margin and in the ice-covered ocean was found to be ca. 2:4:1 (Fig. 1). Our data show a similar trend to that in the study of Carstens et al. (1997). The ice margin offers increased food supply for the foraminifera with higher primary production caused by diatoms that are the major food source for N. pachyderma (sin.) (Hemleben et al. 1989). Diatoms typically do not develop well under permanent ice cover due to the light limitation, but they prefer seasonally stratified water conditions that occur in the marginal ice zone (Smith et al. 1987; Williams 1993). Ice melting increases stratification and consequently the stability of the water column, which triggers phytoplankton blooms (Alexander 1980), and the enhanced phytoplankton biomass along the ice margin may have led to a major increase in absolute abundances of planktic foraminifera.
Vertical distribution
Both N. pachyderma (sin.) and T. quinqueloba had maximum absolute abundances between 0 and 100 m water depth. However, the relative abundances per depth interval (Fig. 4) showed that in the western Fram Strait, at the ice-covered stations more specimens of N. pachyderma (sin.) lived close to the surface (above 100 m), while in the eastern part of the strait the majority of the population was found deeper, between 50 and 200 m. Simstich et al. (2003) analysed plankton and several core-top samples from the Nordic seas, taken mainly south of our transect (80°N–62°N). They reported on a shallow apparent calcification depth in T. quinqueloba over most of the studied profile. If we assume that foraminifera calcify in the water depth where they are most abundant, these results are consistent with our findings. In their study, going from west to the east, the habitat of N. pachyderma (sin.) seems to sink deeper in the Atlantic Water off Norway, similar to what we observe from our samples taken from the warm and saline WSC. These results contrast with the findings of Carstens et al. (1997), who concluded that N. pachyderma (sin.) and T. quinqueloba follow the path of the Atlantic Water submerging below Polar Water and therefore live deeper in the western part of the strait. During our sampling period, in the western Fram Strait, the surface Polar water masses extended down to 200 m. As none of the studies on planktic foraminifera conducted in the same area (including the study of Carstens et al. 1997) revealed an average depth habitat below 200 m, the scenario described by Carstens et al. (1997) seems rather unlikely here. Furthermore, previous investigations revealed that food availability strongly affects the distribution of planktic foraminifera (Schiebel & Hemleben 2005). Thus, the depth habitat of these protozoa might be controlled by the position of the chlorophyll a maximum (Fairbanks & Wiebe 1980; Kohfeld et al. 1996) rather than by the water mass distribution. From the summer months in the Northeast Water Polynya, with oceanographic conditions similar to the western Fram Strait, Kohfeld et al. (1996) reported maximum chlorophyll a concentrations and consequently highest abundances of N. pachyderma (sin.) between 20 and 80 m water depth, which accords with the results presented here. Hirche et al. (1991) also found, for the same period of the year in Fram Strait, chlorophyll a maxima in the open ocean at greater depth than under the ice coverage. Assuming that the depth distribution of N. pachyderma (sin.) is indeed predominantly controlled by food availability in this area, the deeper depth habitat reported by Carstens et al. (1997) could be explained by different sampling periods. It is possible that during the above-mentioned experiments in late summer, the ice coverage was not as intact as in June/July and consequently, due to the higher translucency, the deep chlorophyll maximum might have descended/extended to deeper layers. Planktic foraminifera respond to the redistribution of chlorophyll within several days (Schiebel et al. 2001) and may therefore have followed the descent of the chlorophyll maximum. Another possible explanation for the different findings might be the sampling in different ontogenetic stages. N. pachyderma (sin.) and T. quinqueloba are known to reproduce on a synodic lunar cycle (Bijma et al. 1990; Schiebel & Hemleben 2005) and for reproduction they simultaneously descend from their average depth habitat, possibly to the greatest depth they inhabit during their ontogenetic cycle (Berberich 1996; Schiebel & Hemleben 2005). Volkmann (2000) also assumed that N. pachyderma (sin.) prefers shallower water depths under permanent ice coverage. In her study, this species showed maximum abundances in the narrow temperature layer between −1.5 and −1.8°C, while T. quinqueloba did not show such preference to a particular temperature range. In our study, the maximum abundances of N. pachyderma (sin.) and T. quinqueloba were found between 0 and 100 m water depth at every station and therefore in a very wide temperature range (from −1.7 to 7°C; Fig. 2). We therefore assume that the depth habitat of planktic foraminifera in Fram Strait during June/July is more influenced by the distribution of the food source than by temperature.
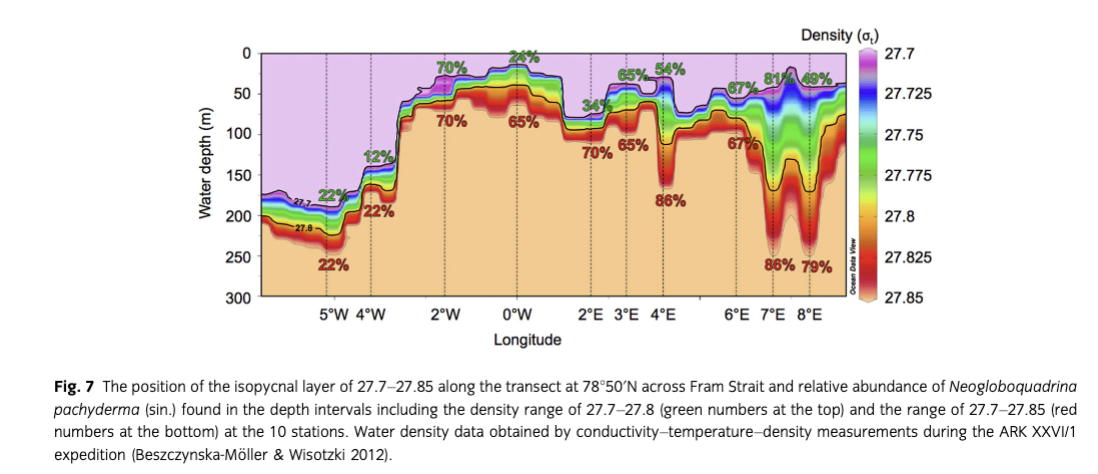
Kozdon et al. (2009) proposed that the average depth habitat of N. pachyderma (sin.) in the Nordic seas is controlled by water density and that this species is bound to a layer between 27.7 and 27.8 (potential density, σt). Along our studied transect during the sampling period, this isopycnal layer was found mainly between 0 and 100 m water depth. There are two exceptions. The first are the stations at 8°E, 7°E and 4°E, where this water mass reaches deeper. The second is the western part of the strait (west of ca. 3°W), on the Greenland shelf, where the whole layer submerged below 150 m. Regarding the relative abundances of N. pachyderma (sin.), our data show that in the eastern and central Fram Strait (2°W–8°E) during the sampling period a broad range from 34 to 81% of the population lived in the depth intervals including the specific density range. Extending the range to 27.85, we found a better correlation with values between 65 and 86% (Fig. 7). In contrast, in the west, in the cold and fresh water masses of the EGC, the depth habitat of N. pachyderma (sin.) was found in significantly shallower water depths than the layer of σt=27.7–27.8. This discrepancy of findings may be related to the nature of the samples and data used. While the water mass and foraminifer data of our study are from just a short time interval and may be representative of only one early summer plankton bloom, they are nonetheless consistent in being obtained at the same time and place (i.e., station and water depth). Kozdon et al. (2009) calculated the calcification depth of N. pachyderma (sin.) using a long-term oceanographic database and temperatures obtained from paired Mg/Ca and δ44/40Ca measurements of shells picked from sediment surface samples. Such samples integrate foraminifer assemblages over the entire plankton growth seasons in a certain number of years, depending on sedimentation rates and bioturbation intensity. Furthermore, variable ages of the surface samples analysed (modern to more than 1000 yr ago; see Simstich et al. 2003) may play a more significant role than anticipated by Kozdon et al. (2009), who claimed that the samples “represent modern oceanographic conditions.” Recent work in the Norwegian Sea and Fram Strait (Sejrup et al. 2010; Spielhagen et al. 2011; Werner et al. 2011) has demonstrated that temperature variations of 2°C and more occurred in the Atlantic Water layer during the last one to two millennia. Particularly in areas with low sedimentation rates (i.e., in ice-covered regions), sediment surface samples may thus rather represent earlier and colder periods (e.g., the Little Ice Age) than just modern, warmer conditions of the comparatively short Industrial Period. For this reason, the isopycnals calculated by Kozdon et al. (2009) on the basis of data extracted from a hydrographic database may not always correspond to the average conditions during deposition of the related sediment surface samples. Our comparison of in situ sampled foraminifera and measurements avoid such potential bias and indicates a wider density range for the habitat of N. pachyderma (sin.).
Comparison with the sedimentary records
Comparison of plankton tow results with those from the sediment surface has important implications for palaeoceanographic reconstructions because in such studies core-top samples are usually assumed to represent the recent foraminiferal fauna and therefore modern conditions. In our study area, the planktic foraminiferal species composition in the overlying water column is fairly well reflected in the sediment surface samples: the polar N. pachyderma (sin.) clearly dominates all the samples, with relative abundances of the subpolar T. quinqueloba varying between 8 and 24%. Again, the differences between plankton tow and sediment data can most likely be ascribed to the early summer sampling of the water column. Lateral advection of foraminifera is not considered a major factor because mean transport distances in Fram Strait are only 25–50 km for N. pachyderma (sin.) and 50–100 km for T. quinqueloba, as determined for a sediment trap at 1125 m water depth (von Gyldenfeldt et al. 2000). In the east and the west of the study area (where differences between plankton tow and sediment data are largest), the flow direction of warm and cold water masses is largely normal to the sampling transect and transport effects of subpolar and polar species on the species assemblages on the sea floor can be expected to be minor. Furthermore, the increase in abundances in the water column at the sea-ice margin compared to the open ocean and under the ice cover is also seen on the sediment surface (Fig. 5). Comparison of the relative abundances of N. pachyderma (sin.) and T. quinqueloba in the depth intervals of 0–50 m and 50–100 m to those from the sediment surface (Fig. 6) suggests that the sediment surface samples relate best to the fauna living between 50 and 100 m water depth. This hypothesis is supported by the fact that in the water column at almost all stations both species were most abundant in this depth interval. The congruence of the fauna found in the water column and on the sediment surface indicates that in this area processes that can influence the deposition of shells (e.g., lateral transport, selective dissolution) do not change significantly the distribution during settling. Since Fram Strait is relatively narrow and interannual west–east variability in the position of the average summer sea-ice margin can be high, effects of fluctuations related to centennial-scale climate changes may be more subdued in the sediment surface samples than in the Nordic seas. As a consequence, the large-scale oceanic regimes are more obvious from the faunal compositions. This suggests that planktic foraminifera in Fram Strait sediments provide a fairly reliable proxy for palaeo-water mass characterization.
Conclusions
In Fram Strait, the distribution pattern of the two dominating species, Neogloboquadrina pachyderma (sin.) and Turborotalita quinqueloba shows a distinct relation to the complex hydrography. Warm and saline Atlantic waters yield higher abundances of the subpolar T. quinqueloba than the cold and fresh waters of the EGC, while the polar N. pachyderma (sin.) shows an exactly reverse tendency.
The ratio between absolute abundances of planktic foraminifera in the open ocean, at the ice margin and in the ice-covered ocean is ca. 2:4:1. High abundances in the marginal ice zone are related to the high primary production triggered by the strongly stratified water column at the ice margin.
The depth habitats of N. pachyderma (sin.) and T. quinqueloba in the area seem predominantly controlled by the availability of food. Both species dwell shallower under the ice cover than under ice-free conditions, possibly following the position of the deep chlorophyll maximum. The effect of other water column properties, like temperature, on the depth habitat seems to be masked. We could not verify the hypothesis of Kozdon et al. (2009), which associates the vertical distribution of N. pachyderma (sin.) to a discrete isopycnal band. This discrepancy might be due to various factors, for example, differences in applied methods and palaeoceanographic variability in the study area in the recent past.
Sediment surface samples, often used for palaeoceanographic reconstructions, fairly well reflect the composition of the foraminiferal fauna living in the overlying water column. In Fram Strait, the species composition and relative abundance in the sediments correlate best with that of the fauna living between 50 and 100 m water depth. In this area—and probably beyond—planktic foraminifera may therefore be used as proxies for subsurface water conditions.
Acknowledgements
The authors wish to thank the captain and crew of the RV Polarstern and the Alfred Wegener Institute, Helmholtz Centre for Polar and Marine Research in Bremerhaven for support during ARK XXVI/1 cruise. We are indebted to Dr Angelina Kraft for assisting in the deployments of the MultiNet sampler and to Maciej Telesinski for collecting the sediment surface samples for this study. Dr Agnieszka Beszczynska-Möller and Andreas Wisotzki provided the CTD data for this study. Thanks are due to Dr Hartmut Schulz for kindly providing the low temperature vacuum asher at the University of Tübingen and for his hospitality and technical assistance during the process. Thanks also go to Dr Dorothea Bauch, who helped during the preparation phase for the cruise and for her valuable comments and suggestions throughout the study, and to Dr Daniele de Corte for reviewing the manuscript. We thank Thomas M. Cronin and one anonymous reviewer for their constructive feedback and insightful comments. This research was conducted with funding of the European Community’s 7th Framework Programme FP7 2007/2013, Marie-Curie Actions, under Grant Agreement no. 238111–CASE ITN. RFS thanks the Academy of Sciences, Humanities and Literature Mainz for financial support through the Akademienprogramm.
References
- Alexander V. 1980. Interrelationships between the seasonal sea ice and biological regimes. Cold Regions Science and Technology 2, 157–178. Publisher Full Text
- Andrijanic S. 1988. Geographical distribution of living planktonic foraminifera (Protozoa) off the east coast of Australia. Australian Journal of Marine and Freshwater Research 39, 71–85. Publisher Full Text
- Bé A.W.H. 1960. Some observations on Arctic planktonic foraminifera. Contributions from the Cushman Foundation for Foraminiferal Research 11, 64–68.
- Bé A.W.H. & Hutson W.H. 1977. Ecology of planktonic foraminifera and biogeographic patterns of live and fossil assemblages in the Indian Ocean. Micropaleontology 23, 369–414. Publisher Full Text
- Berberich D. 1996. Die planktische Foraminifere Neogloboquadrina pachyderma (Ehrenberg) im Weddellmeer, Antarktis. (The planktic foraminifer Neogloboquadrina pachyderma (Ehrenberg) in the Weddell Sea, Antarctica.) Berichte zur Polarforschung 195. Bremerhaven: Alfred Wegener Institute.
- Beszczynska-Möller A. & Wisotzki A. 2012. Physical oceanography during POLARSTERN cruise ARKXXVI/1. Bremerhaven: Helmholtz Centre for Polar and Marine Research, Alfred Wegener Institute.
- Bijma J., Erez J. & Hemleben C. 1990. Lunar and semi-lunar reproductive cycles in some spinose planktonic foraminifers. Journal of Foraminiferal Research 20, 117–127. Publisher Full Text
- Carstens J., Hebbeln D. & Wefer G. 1997. Distribution of planktic foraminifera at the ice margin in the Arctic (Fram Strait). Marine Micropaleontology 29, 257–269. Publisher Full Text
- Carstens J. & Wefer G. 1992. Recent distribution of planktonic foraminifera in the Nansen Basin, Arctic Ocean. Deep-Sea Research Part I 39, 507–524. Publisher Full Text
- Dickson R.R., Osborn T.J., Hurrell J.W., Meincke J., Blindheim J., Adlandsvik B., Vinje T., Alekseev G. & Maslowski W. 2000. The Arctic Ocean response to the North Atlantic Oscillation. Journal of Climate 13, 2671–2696. Publisher Full Text
- Ehrenberg C.G. 1861. Über die Tiefgrund-Verhältnisse des Oceans am Eingange der Davisstrasse und bei Island. (About sea bottom conditions at the mouth of Davis Strait and at Iceland.) Monatsbericht der Königlichen Preussischen Akademie der Wissenschaften zu Berlin. Pp. 275–315.
- Fairbanks R.G. & Wiebe P.H. 1980. Foraminifera and chlorophyll maximum: vertical distribution, seasonal succession, and palaeoceanographic significance. Science 209, 1524–1525. Publisher Full Text
- Hansen A. & Knudsen K.L. 1995. Recent foraminiferal distribution in Freemansundet and Early Holocene stratigraphy on Edgeøya, Svalbard. Polar Research 14, 215–238. Publisher Full Text
- Hemleben C., Spindler M. & Anderson O.R. 1989. Modern planktonic foraminifera. New York: Springer.
- Hirche H.-J., Baumann M.E.M., Kattner G. & Gradinger R. 1991. Plankton distribution and the impact of copepod grazing on primary production in Fram Strait, Greenland Sea. Journal of Marine Systems 2, 477–494. Publisher Full Text
- Jakobsson M., Mayer L., Coakley B., Dowdeswell J.A., Forbes S., Fridman B., Hodnesdal H., Noormets R., Pedersen R., Rebesco M., Schenke H.W., Zarayskaya Y., Accettella D., Armstrong A., Anderson R.M., Bienhoff P., Camerlenghi A., Church I., Edwards M., Gardner J.V., Hall J.K., Hell B., Hestvik O., Kristoffersen Y., Marcussen C., Mohammad R., Mosher D., Nghiem S.V., Pedrosa M.T., Travaglini P.G. & Weatherall P. 2012. The International Bathymetric Chart of the Arctic Ocean (IBCAO) version 3.0. Geophysical Research Letters 39, L12609, doi: 10.1029/2012GL052219. Publisher Full Text
- Johannessen O.M. 1986. Brief overview of the physical oceanography. In B.G. Hurdle (ed.): The Nordic seas. Pp. 103–127. Springer: New York.
- Jones E.P. 2001. Circulation in the Arctic Ocean. Polar Research 20, 139–146. Publisher Full Text
- Jonkers L., Brummer G.-J.A., Peeters F.J.C., van Aken H.M. & De Jong M.F. 2010. Seasonal stratification, shell flux, and oxygen isotope dynamics of left-coiling N. pachyderma (sin.) and T. quinqueloba in the western subpolar North Atlantic. Paleoceanography 25, PA2204, doi: 10.1029/2009PA001849. Publisher Full Text
- Kohfeld K.E., Fairbanks R.G., Smith S.L. & Walsh I.D. 1996. Neogloboquadrina pachyderma (sinistral coiling) as palaeoceanographic tracers in polar oceans: evidence from Northeast Water Polynya plankton tows, sediment traps, and surface sediments. Paleoceanography 11, 679–699. Publisher Full Text
- Kozdon R., Eisenhauer A., Weinelt M., Meland M.Y. & Nürnberg D. 2009. Reassessing Mg/Ca temperature calibrations of Neogloboquadrina pachyderma (sinistral) using paired Ca and Mg/Ca measurements. Geochemistry, Geophysics, Geosystems 10, Q03005, doi: 10.1029/2008GC002169. Publisher Full Text
- Kuroyanagi A. & Kawahata H. 2004. Vertical distribution of living planktonic foraminifera in the seas around Japan. Marine Micropaleontology 53, 173–196. Publisher Full Text
- Naomi T.S., Mathew K.J. & Antony G. 1990. Studies on the distribution of recent planktonic foraminifera in the Arabian Sea and the Bay of Bengal. In K.J. Mathew (ed.): Proceedings of the First Workshop on Scientific Results of FORV Sagar Sampada, Cochin, 5–7 June 1989. Pp. 71–79. Cochin, India: Central Marine Fisheries Research Institute.
- Natland M.L. 1938. New species of foraminifera from off the west coast of North America and from the later Tertiary of the Los Angeles Basin. Scripps Institute of Oceanography Bulletin 4, 137–164.
- Rudels B., Friedrich H.J. & Quadfasel D. 1999. The Arctic Circumpolar Boundary Current. Deep-Sea Research Part II 46, 1023–1062. Publisher Full Text
- Rudels B., Meyer R., Fahrbach E., Ivanov V.V., Østerhus S., Quadfasel D., Schauer U., Tverberg V. & Woodgate R.A. 2000. Water mass distribution in Fram Strait and over the Yermak Plateau in summer 1997. Annales Geophysicae 18, 687–705. Publisher Full Text
- Schiebel R. & Hemleben C. 2000. Interannual variability of planktic foraminiferal populations and test flux in the eastern North Atlantic Ocean (JGOFS). Deep-Sea Research Part II 47, 1809–1852. Publisher Full Text
- Schiebel R. & Hemleben C. 2005. Modern planktic foraminifera. Paläontologische Zeitschrift 79, 135–148.
- Schiebel R., Waniek J., Bork M. & Hemleben C. 2001. Planktic foraminiferal production stimulated by chlorophyll redistribution and entrainment of nutrients. Deep-Sea Research Part I 48, 721–740. Publisher Full Text
- Sejrup H.P., Lehman S.J., Haflidason H., Noone D., Muscheler R., Berstad I.M. & Andrews J.T. 2010. Response of Norwegian Sea temperature to solar forcing since 1000 A.D. Journal of Geophysical Research—Oceans 115, C12034, doi: 10.1029/2010JC006264. Publisher Full Text
- Simstich J., Sarnthein M. & Erlenkeuser H. 2003. Paired δ18O signals of Neogloboquadrina pachyderma (s) and Turborotalita quinqueloba show thermal stratification structure in the Nordic seas. Marine Micropaleontology 48, 107–125. Publisher Full Text
- Smith W.O., Jr., Baumann M.E.M., Wilson D.L. & Aletsee L. 1987. Phytoplankton biomass and productivity in the marginal ice zone of the Fram Strait during summer 1984. Journal of Geophysical Research—Oceans 92, 6777–6786. Publisher Full Text
- Spielhagen R.F., Werner K., Sorensen S.A., Zamelczyk K., Kandiano E., Budeus G., Husum K., Marchitto T.M. & Hald M. 2011. Enhanced modern heat transfer to the Arctic by warm Atlantic water. Science 331, 450–453. Publisher Full Text
- Vinje T. 2001. Fram Strait ice fluxes and atmospheric circulation: 1950–2000. Journal of Climate 14, 3508–3517. Publisher Full Text
- Vinje T.E. 1977. Sea ice conditions in the European sector of the marginal seas of the Arctic, 1966–75. Norsk Polarinstitutt Årbok 1975, 163–174.
- Volkmann R. 2000. Planktic foraminifers in the outer Laptev Sea and the Fram Strait—modern distribution and ecology. Journal of Foraminiferal Research 30, 157–176. Publisher Full Text
- von Gyldenfeldt A.-B., Carstens J. & Meincke J. 2000. Estimation of the catchment area of a sediment trap by means of current meters and foraminiferal tests. Deep-Sea Research Part II 47, 1701–1717. Publisher Full Text
- Werner K., Spielhagen F.R., Bauch D., Hass H.C., Kandiano E. & Zamelczyk K. 2011. Atlantic Water advection to the eastern Fram Strait—multiproxy evidence for late Holocene variability. Paleogeography, Paleoclimatology, Paleoecology 308, 264–276. Publisher Full Text
- Williams K.M. 1993. Ice sheet and ocean interactions, margin of the East Greenland ice sheet (14 Ka to present): diatom evidence. Paleoceanography 8, 69–83. Publisher Full Text
Related Articles
Ruibo Lei
Petra Heil
Jia Wang
Zhanhai Zhang
et al.
Eva-Maria Nöthig
Astrid Bracher
Anja Engel
Katja Metfies
et al.
Alexander Vetrov
Evgeny Romankevich
Evgeny Gusev
Pavel Rekant
Valery Kaminsky
Alexey Krylov
et al.