This article is published under a Creative Commons license and not by the author of the article. So if you find any inaccuracies, you can correct them by updating the article.

The effect of an experimental decrease in salinity on the viability of the Subarctic planktonic foraminifera Neogloboquadrina incompta



Mattia Greco
Tine L. Rasmussen
Michal Kucera
Julie Meilland
Kasia Zamelczyk
Published: Aug. 27, 2020
Latest article update: Aug. 3, 2023
This article is published under the license




Abstract
Chemical signatures in the calcite of shells of polar and subpolar planktonic foraminifera have been frequently used to trace and quantify past meltwater discharge events. This approach assumes that the foraminifera can tolerate low salinity under extended periods. To obtain a first experimental constraint on salinity tolerance of Subarctic foraminifera, we carried out a culturing experiment with specimens of the subpolar species Neogloboquadrina incompta collected in the northern Norwegian Sea off Tromsø in October 2018. The foraminifera were exposed to a gradient of salinities between 35 and 25 PSU. Survival was monitored over 26 days by measuring the extent of the rhizopodial network. Although chamber growth only occurred in one of the observed specimens, likely due to the largely unknown dietary preference of the species, we observed a strong differential rhizopodial activity pattern along the gradient. The highest rhizopodial activity occurred at salinity between 35 and 31 PSU. The species is clearly able to survive long-term exposure to salinities as low as 28, but no rhizopodial activity and signs of cytoplasm degradation were observed in all specimens exposed to 25 PSU. These preliminary observations provide the first direct evidence for the salinity tolerance of N. incompta, indicating a range of salinity that could be plausibly expected to be recorded in the chemistry of fossil shells of the species.
Keywords
Planktonic foraminifera, salinity, laboratory observations, meltwater events
Introduction
The chemical and isotopic composition of fossil shells of planktonic foraminifera is a well-established approach to investigate the past state of the ocean (e.g., Ravelo & Hillaire-Marcel 2007; Pearson 2012). For example, the oxygen isotopic signature (δ18O) in shells of Neogloboquadrina incompta has been used to infer the presence of meltwater injected into the surface ocean by icebergs (McManus et al. 1999; Came et al. 2007; Rashid & Boyle 2007; Voelker et al. 2009). The reconstructions are based on the assumption that the calcification of the shell and, therefore, the incorporation of the chemical signal occurred within the water layer affected by the discharged meltwater. This is particularly relevant in situations where the properties of the target water layer may be modified to a degree that is too hostile for the survival of the foraminifera. In this scenario, specimens of the species could be largely excluded from surface low-salinities habitat, and the oxygen-isotope composition of the remaining specimens dwelling deeper would be recording conditions below the meltwater layer, leading to a systematic underestimation of the surface salinity anomaly. Indeed, past meltwater injections in the North Atlantic likely had a magnitude sufficient to modify surface salinity below the range of naturally occurring values in the present ocean (Hemming 2004). Among the species of planktonic foraminifera occurring in the North Atlantic during these events, especially in the more distal part of the iceberg discharge plume, is N. incompta (Dickson et al. 2008; Voelker et al. 2009).
However, to date, no experimental data are available to constrain the range of salinities under which N. incompta survives and which it thus could potentially record.
Most existing experiments in which planktonic foraminifera were exposed to a gradient of environmental parameters have been carried out on tropical to temperate species (McCrea 1950; Bé et al. 1977; Bijma et al. 1990; Lea et al. 1999; Davis et al. 2017; Bertlich et al. 2018; Fehrenbacher et al. 2018; LeKieffre et al. 2018). High-latitude planktonic foraminifera have been rarely kept in culture (Manno et al. 2012), and standardized culturing protocols have not been established for the cultivation of these species under cold conditions (Kozdon et al. 2009; Schiebel et al. 2018). Here, we present the results from a preliminary laboratory experiment on the Subarctic planktonic foraminifera N. incompta with the purpose to constrain the salinity tolerance of the species. With this experiment, we aim to provide a first insight into the changes in the physiology and viability of N. incompta in response to different salinity conditions and introduce a novel way of monitoring its physiology, applicable in the absence of growth, by measuring the extent of its rhizopodial network.
Materials and methods
Sampling
The experiment and the microscope observations were performed in a cold room in one of the facilities of UiT—The Arctic University of Norway in Tromsø.
Specimens of N. incompta were collected during a cruise on the RV Helmer Hansen in October 2018 to the shelf and slope of northern Norway off Tromsø, specifically an area of the shelf known as Håkjerringdjupet. In the sampling area, surface water temperature ranged between 6 and 10°C. The encountered community of planktonic foraminifera was dominated by N. incompta, which gave us the opportunity to study the salinity tolerance of this species at the lower end of its thermal range, under conditions that can be expected to resemble those of past meltwater injections, with cold temperatures due to iceberg melting. Specimens were sampled from a water depth between 0 and 100 m, using a WP2 plankton net (63 µm mesh size) that was towed vertically. The retrieved specimens were picked on board and incubated in jars containing seawater at 35 PSU previously filtered through a 0.22 µm nitrate cellulose filter (Fig. 1) and allowed to recover for ca. 16 hr at treatment temperature (6°C).

Culture methods
Onshore, cytoplasm-bearing N. incompta specimens were transferred from the collection flasks into Petri dishes and, after six hours, a fraction of the specimens was transferred into new Petri dishes with a salinity lowered by 3–4 PSU to avoid osmotic shock. This acclimatization procedure was repeated from the Petri dish with lowered salinity at intervals of six hours until the minimum tested salinity of 25 PSU was reached. The tested range of salinities was chosen to reach below 30 PSU, which is the lower limit of salinity estimates in the Heinrich meltwater layers (Maslin et al. 1995; De Vernal & Hillaire-Marcel 2000). The culturing medium for the treatments (35 PSU/Control, 31 PSU, 28 PSU and 25 PSU) was obtained by consecutive dilutions of ambient seawater with MilliQ water. Salinity was measured by means of a digital refractometer. From the treatment series, cytoplasm-bearing specimens of N. incompta were removed and cultured individually under the treatment salinity in 75 ml Falcon flasks and constant temperature of 6°C in a cold room under eight hours light cycles (intensity of 150 µmol photons m–2 s–1 [Manno et al. 2012]). They were fed daily with 30 µl autoclaved marine microalgae Nannochloropsis food mix (30 µl Nannochloropsis concentrate: 200 ml filtered seawater), attempting to simulate a diet involving phytoplankton detritus. A population of 16 specimens in the size range of 95–203 µm was initially selected for the experiment. A larger population number was not possible with the given sampled population size and the effort associated with individual monitoring. After the introduction of the treatment gradient, one specimen was left for the individual culturing in the control (ambient) treatment (35 PSU), three for 31 PSU, three for 28 PSU and two for 25 PSU.
Analyses
The response of the individual specimens to the treatment was monitored until cytoplasm decay was observed (Fig. 2d). Cytoplasm-bearing specimens that did not display any rhizopodial net for 18 days from the start of the experiment were re-checked after day 22. The foraminifera were photographed using a digital camera attached to an inverted microscope, and the state (colour) of the cytoplasm was reported. The software ImageJ, Version 1.8.0 (Schneider et al. 2012), was used to measure the rhizopodial activity of each specimen calculated as the ratio between the maximum shell diameter and the maximum extension of the rhizopods (Fig. 3). This parameter was chosen because both measurements are largely invariant to rotation on a plane (the specimens were not floating during observations) and because estimating the number of extended rhizopodial is difficult and more ambiguous than a determination of the maximum extension length. Repeated measurements on selected specimens indicate that the uncertainty on the determination of the maximum shell diameter is 3%, and assuming similar uncertainty on the rhizopodial extension, the resulting uncertainty on the index should be about 6%. After the experiment, the cultured specimens were photographed using SEM at the University of Bremen.
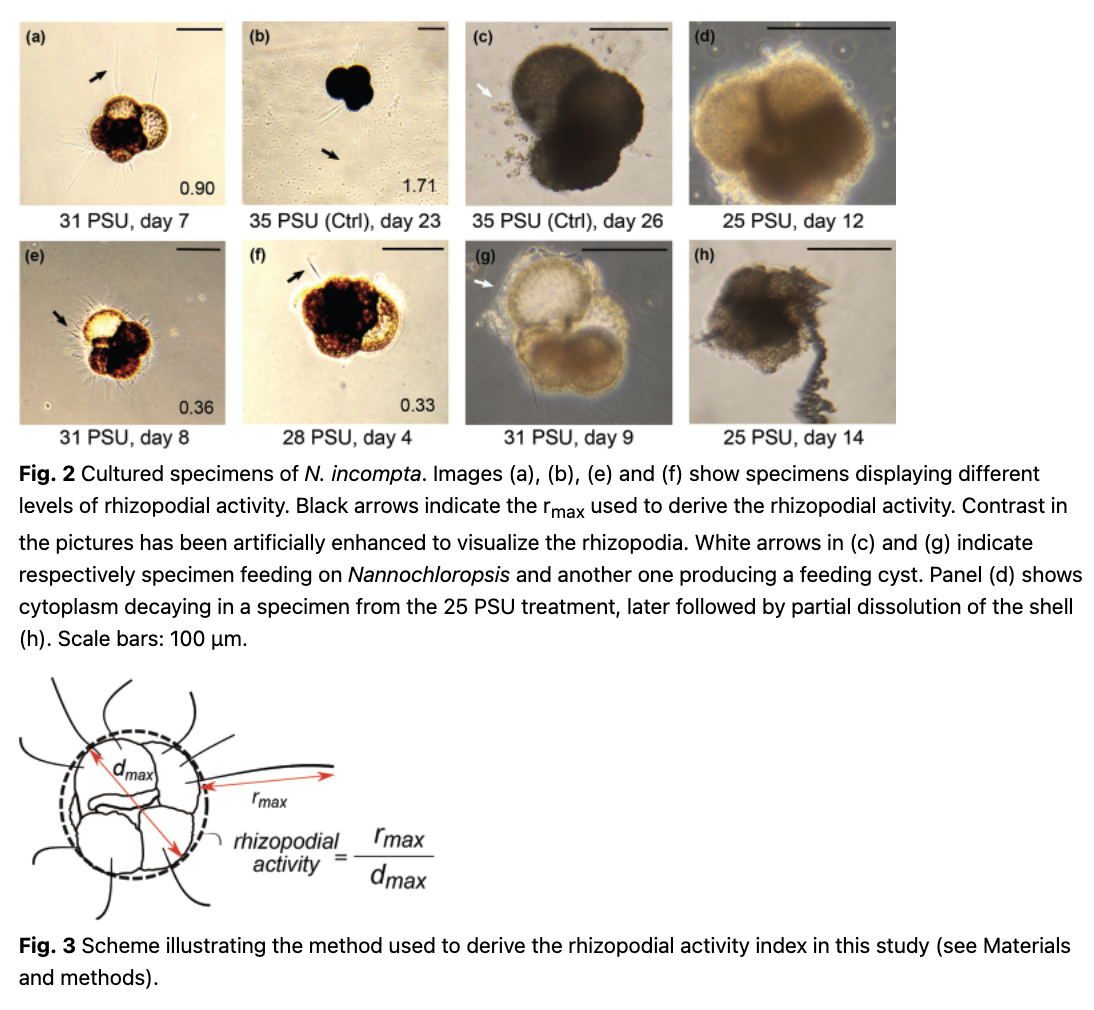
Given the small scale of our experiment, we decided to refrain from statistical analyses.
Results
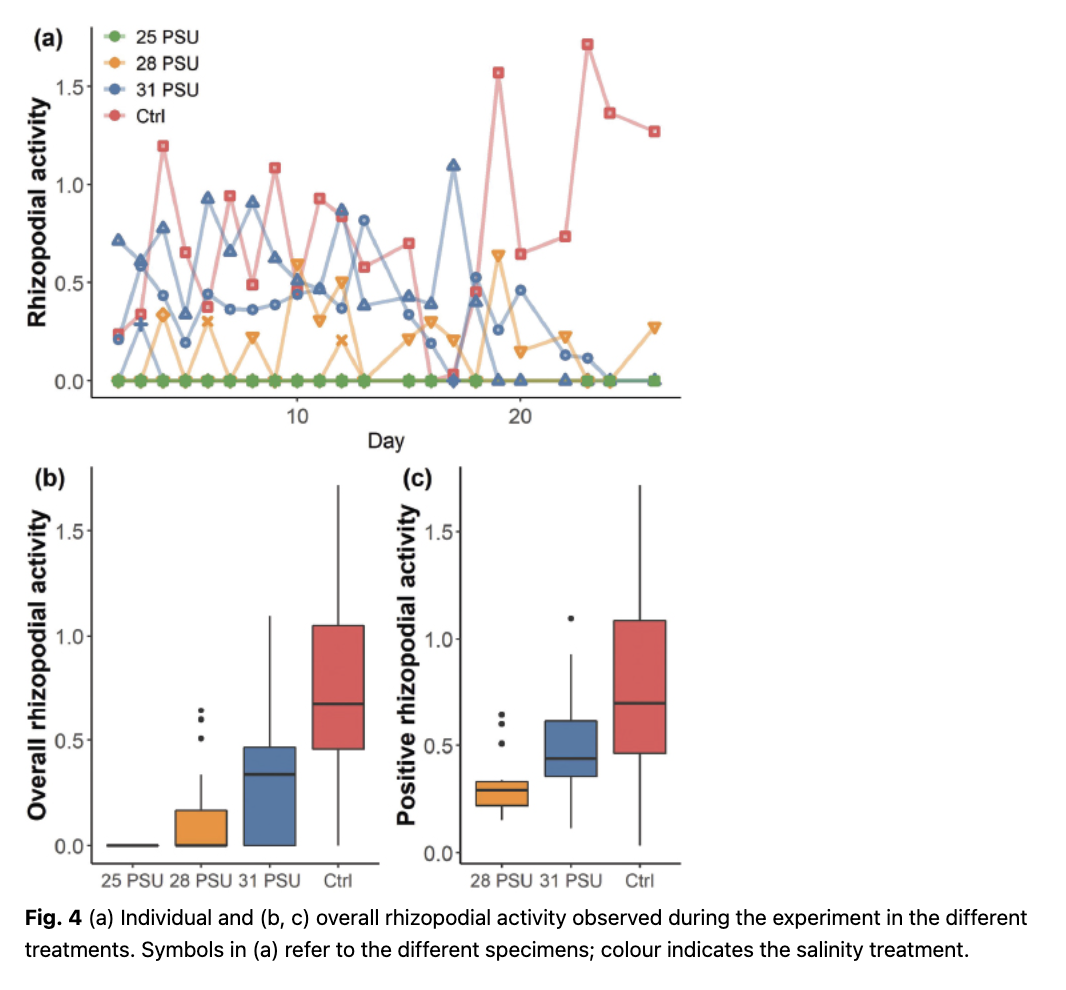
In the control treatment, the viable N. incompta specimen survived for the entire duration of the experiment, displaying the highest rhizopodial activity registered (1.71; Fig. 4). In the 31 PSU treatment, one specimen survived until day 3, formed a kummerform final chamber and showing signs of shell thickening (Fig. 4). The remaining two specimens showed rhizopodial activity until days 18 and 24, respectively. The overall average rhizopodial activity was lower in this treatment than in the control (Fig. 4b). The same applies to the average activity for all observations when extended rhizopodia were observed (Fig. 4c). At the 28 PSU, two of the specimens stopped displaying rhizopodial activity after day 12 and later showed signs of cytoplasm decay. Only one specimen survived until the end of the experiment. The overall average rhizopodial activity, as well as the average activity for all observations when extended rhizopodia were observed, was the lowest (Fig. 4). None of the specimens cultured at 25 PSU showed rhizopodial activity during the experiment and both specimens showed signs of cytoplasm decay after day 15 (Fig. 2d).
Discussion
Our observations indicate that N. incompta rhizopodial activity decreases on exposure to salinity from 35 to 28 PSU, but survival under an extended period of time (weeks) is possible within this salinity range, whereas it appears that extended exposure to 25 PSU is lethal. There are no earlier experimental observations on the salinity tolerance of this species, and ambient salinities in the modern ocean where planktonic foraminifera occur, even in the Arctic where the lowest salinity conditions are expected, are always >29 PSU (Greco et al. 2019). However, Bijma et al. (1990) presented data on salinity limits of the related species Neogloboquadrina dutertrei. Although these authors used a different definition of viability based on growth, they observed that the vital processes of the tested specimens of N. dutertrei were completely inhibited at 25 PSU. This observation agrees with our results on N. incompta. It is important to note that the ability of N. incompta to survive under reduced salinities under laboratory conditions does not necessarily mean that it will inhabit a similarly low-saline meltwater lens in the natural environment. Indeed, laboratory experiments can only constrain the maximum range of salinities under which survival in the field may occur.
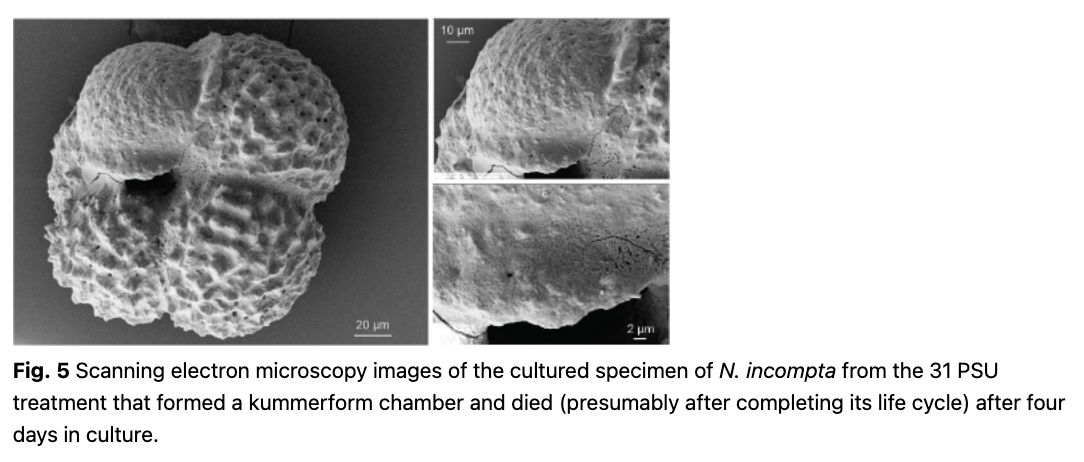
In the present experiments, one of the cultured specimens showed signs of chamber formation and thickening under the light microscope. As no calcification label was added to the culture seawater, we confirmed the observation by subsequent analyses of the recovered shell using SEM. This revealed the addition of a kummerform chamber and of shell-thickening by secondary calcification (Fig. 5). Both observations are consistent with the normal behaviour prior to gametogenesis in planktonic foraminifera (Hemleben et al. 1989). This indicates that the laboratory conditions in our experiment did not preclude growth or calcification and, therefore, the termination of its natural life cycle explains why this specimen died so early despite exposure to the non-lethal salinity level of 31 PSU. In the light of this observation, it remains unclear why the remaining specimens in our experiment survived but did not grow.
A possible explanation may lie in the low cultivation temperature of 6°C. Indeed, the few previous culturing studies on N. incompta grown under different temperatures reported no growth in specimens cultured at 6°C, but growth occurred at 9°C (Von Langen et al. 2005; Davis et al. 2017). Unfortunately, both culturing studies were carried out in the Pacific, which is inhabited by a different cryptic species of N. incompta (Darling et al. 2006), making it difficult to directly transfer these observations on the North Atlantic species. Alternatively, it could be that the autoclaved Nannochloropsis used for feeding the cultured N. incompta does not represent a suitable food source for this species. In the previous experiments, the cultured N. incompta specimens were fed with freshly killed Artemia (Von Langen et al. 2005; Davis et al. 2017), but recent molecular investigations revealed that this species may feed on bacteria (Bird et al. 2018). With the food preference of this species is unknown (Artemia cannot be the natural prey and has been taken as a substitute for marine copepods), we opted for autoclaved Nannochloropsis, assuming that it emulates the likely available food found below the sunlit layer (N. incompta is a subsurface species [Rebotim et al. 2017]) and considering that it was found to be accepted by other foraminifera (Schmidt et al. 2015). We observed that the autoclaved Nannochloropsis was accepted by N. incompta and collected by its rhizopodial network (Fig. 2c, g), forming a feeding cyst (Spindler et al. 1984; Hemleben et al. 1989; Heinz et al. 2005; Bird et al. 2018), but it is possible that either the quantity or quality of the food was insufficient to facilitate shell growth.
Conclusions
Our study provides first experimental and preliminary evidence for physiological stress in N. incompta with decreasing salinity under “polar” conditions. We show that the species survives extended chronic exposure from 35 to 28 PSU, and we interpret the complete absence of extended rhizopods at 25 PSU as evidence for physiologically lethal conditions. Our experiment indicates that quantification of the extent of rhizopodial activity may be an effective measure of physiological health, which can be used even in situations and at timescales where no shell growth occurs. Because of the small number of specimens investigated, these conclusions require validation by further experiments, but the preliminary results provide a context for assessing the salinity tolerance of this species and can serve as a basis to better interpret the palaeoclimatic reconstructions based on fossil shells of N. incompta.
Data availability
Data in support of the findings are available on figshare at https://doi.org/10.6084/m9.figshare.11309627.v1.
Acknowledgements
The captain and crew of the RV Helmer Hansen and engineers B.R. Olsen and T. Holm are gratefully acknowledged for their support of the work during the sample collection. We also thank the Department of Geosciences at UiT—The Arctic University of Norway, Tromsø, for providing logistical support.
References
- Bé A.W.H., Hemleben C., Anderson O.R., Spindler M., Hacunda J. & Tuntivate-Choy S. 1977. Laboratory and field observations of living planktonic foraminifera. Micropaleontology 23, 155–179, doi: 10.2307/1485330.
- Bertlich J., Nürnberg D., Hathorne E.C., de Nooijer L.J., Mezger E.M., Kienast M., Nordhausen S., Reichart G., Schönfeld J. & Bijma J. 2018. Salinity control on Na incorporation into calcite tests of the planktonic foraminifera Trilobatus sacculifer—evidence from culture experiments and surface sediments. Biogeosciences 15, 5991–6018, doi: 10.5194/bg-15-5991-2018.
- Bijma J., Faber W.W. & Hemleben C. 1990. Temperature and salinity limits for growth and survival of some planktonic foraminifers in laboratory cultures. Journal of Foraminiferal Research 20, 95–116, doi: 10.2113/gsjfr.20.2.95.
- Bird C., Darling K.F., Russell A.D., Fehrenbacher J.S., Davis C.V., Free A. & Ngwenya B.T. 2018. 16S rRNA gene metabarcoding and TEM reveals different ecological strategies within the genus Neogloboquadrina (planktonic foraminifer). PLoS One 13, 83–95, doi: 10.1371/journal.pone.0191653.
- Came R.E., Oppo D.W. & McManus J.F. 2007. Amplitude and timing of temperature and salinity variability in the subpolar North Atlantic over the past 10 k.y. Geology 35, 315–318, doi: 10.1130/G23455A.1.
- Darling K.F., Kucera M., Kroon D. & Wade C.M. 2006. A resolution for the coiling direction paradox in Neogloboquadrina pachyderma. Paleoceanography 21, PA2011, doi: 10.1029/2005PA001189.
- Davis C.V., Fehrenbacher J.S., Hill T.M., Russell A.D. & Spero H.J. 2017. Relationships between temperature, pH, and crusting on Mg/Ca ratios in laboratory‐grown Neogloboquadrina foraminifera. Paleoceanography 32, 1137–1152, doi: 10.1002/2017PA003111.
- De Vernal A. & Hillaire-Marcel C. 2000. Sea-ice cover, sea-surface salinity and halo-/thermocline structure of the northwest North Atlantic: modern versus full glacial conditions. Quaternary Science Reviews 19, 65–85, doi: 10.1016/S0277-3791(99)00055-4.
- Dickson A.J., Austin W.E.N., Hall I.R., Maslin M.A. & Kucera M. 2008. Centennial‐scale evolution of Dansgaard‐Oeschger events in the northeast Atlantic Ocean between 39.5 and 56.5 ka B.P. Paleoceanography 23, PA3206, doi: 10.1029/2008PA001595.
- Fehrenbacher J.S., Russell A.D., Davis C.V., Spero H.J., Chu E. & Hönisch B. 2018. Ba/Ca ratios in the non-spinose planktic foraminifer Neogloboquadrina dutertrei: evidence for an organic aggregate microhabitat. Geochimica et Cosmochimica Acta 236, 361–372, doi: 10.1016/j.gca.2018.03.008.
- Greco M., Jonkers L., Kretschmer K., Bijma J. & Kucera M. 2019. Depth habitat of the planktonic foraminifera Neogloboquadrina pachyderma in the northern high latitudes explained by sea-ice and chlorophyll concentrations. Biogeosciences 16, 3425–3437, doi: 10.5194/bg-16-3425-2019.
- Heinz P., Geslin E. & Hemleben C. 2005. Laboratory observations of benthic foraminiferal cysts. Marine Biology Research 1, 149–159, doi: 10.1080/17451000510019114.
- Hemleben C., Spindler M. & Anderson O.R. 1989. Modern planktonic foraminifera. New York: Springer.
- Hemming S.R. 2004. Heinrich events: massive late Pleistocene detritus layers of the North Atlantic and their global climate imprint. Reviews of Geophysics 42, RG1005, doi: 10.1029/2003RG000128.
- Kozdon R., Ushikubo T., Kita N.T., Spicuzza M. & Valley J. W. 2009. Intratest oxygen isotope variability in the planktonic foraminifer N. pachyderma: real vs. apparent vital effects by ion microprobe. Chemical Geology 258, 327–337, doi: 10.1016/j.chemgeo.2008.10.032.
- Lea D.W., Mashiotta T.A. & Spero H.J. 1999. Controls on magnesium and strontium uptake in planktonic foraminifera determined by live culturing. Geochimica et Cosmochimica Acta 63, 2369–2379, doi: 10.1016/S0016-7037(99)00197-0.
- LeKieffre C., Spero H.J., Russell A.D., Fehrenbacher J.S., Geslin E. & Meibom A. 2018. Assimilation, translocation, and utilization of carbon between photosynthetic symbiotic dinoflagellates and their planktic foraminifera host. Marine Biology 165, 104, doi: 10.1007/s00227-018-3362-7.
- Manno C., Morata N. & Bellerby R. 2012. Effect of ocean acidification and temperature increase on the planktonic foraminifer Neogloboquadrina pachyderma (sinistral). Polar Biology 35, 1311–1319, doi: 10.1007/s00300-012-1174-7.
- Maslin M.A., Shackleton N.J. & Pflaumann U. 1995. Surface water temperature, salinity, and density changes in the northeast Atlantic during the last 45,000 years: Heinrich events, deep water formation, and climatic rebounds. Paleoceanography 10, 527–544, doi: 10.1029/94PA03040.
- McCrea J.M. 1950. On the isotopic chemistry of carbonates and a paleotemperature scale. The Journal of Chemical Physics 18, 849–857, doi: 10.1063/1.1747785.
- McManus J.F., Oppo D.W. & Cullen J. L. 1999. A 0.5-million-year record of millennial-scale climate variability in the North Atlantic. Science 283, 971–975, doi: 10.1126/science.283.5404.971.
- Pearson P.N. 2012. Oxygen isotopes in foraminifera: overview and historical review. The Paleontological Society Papers 18, 1–38, doi: 10.1017/S1089332600002539.
- Rashid H. & Boyle E.A. 2007. Mixed-layer deepening during Heinrich events: a multi-planktonic foraminiferal δ18O approach. Science 318, 439–441, doi: 10.1126/science.1146138.
- Ravelo A.C. & Hillaire-Marcel C. 2007. The use of oxygen and carbon isotopes of foraminifera in paleoceanography. In C. Hillaire-Marcel & A. De Vernal (eds): Proxies in Late Cenozoic paleoceanography. Vol. 1. Pp. 735–764. Amsterdam: Elsevier.
- Rebotim A., Voelker A.H.L., Jonkers L., Waniek J.J., Meggers H., Schiebel R., Fraile I., Schulz M. & Kucera M. 2017. Factors controlling the depth habitat of planktonic foraminifera in the subtropical eastern North Atlantic. Biogeosciences 14, 827–859, doi: 10.5194/bg-14-827-2017.
- Schiebel R., Smart S.M., Jentzen A., Jonkers L., Morard R., Meilland J., Michel E., Coxall H.K., Hull P.M., de Garibdel-Thoron T., Aze T., Quillévéré F., Ren H., Sigman D.M., Vonhof H.B., Martínez-García A., Kucera M., Bijma J., Spero H.J. & Haug G.H. 2018. Advances in planktonic foraminifer research: new perspectives for paleoceanography. Revue de Micropaléontologie 61, 113–138, doi: 10.1016/j.revmic.2018.10.00.
- Schmidt C., Morard R., Almogi-Labin A., Weinmann A.E., Titelboim D., Abramovich S. & Kucera M. 2015. Recent invasion of the symbiont-bearing foraminifera pararotalia into the eastern mediterranean facilitated by the ongoing warming trend. PLoS One 10, e0132917, doi: 10.1371/journal.pone.0132917.
- Schneider C.A., Rasband W.S. & Eliceiri K.W. 2012. NIH Image to ImageJ: 25 years of image analysis. Nature Methods 9, 671–675, doi: 10.1038/nmeth.2089.
- Spindler M., Hemleben C., Salomons J.B. & Smit L.P. 1984. Feeding behavior of some planktonic foraminifers in laboratory cultures. Journal of Foraminiferal Research 14, 237–249, doi: 10.2113/gsjfr.14.4.237.
- Voelker A.H.L., De Abreu L., Schönfeld J., Erlenkeuser H. & Abrantes F. 2009. Hydrographic conditions along the western Iberian margin during marine isotope stage 2. Geochemistry, Geophysics, Geosystems 10, Q12U08, doi: 10.1029/2009GC002605.
- von Langen P.J., Pak D.K., Spero H.J. & Lea D.W. 2005. Effects of temperature on Mg/Ca in neogloboquadrinid shells determined by live culturing. Geochemistry, Geophysics Geosystems 6, Q10P03, doi: 10.1029/2005GC000989.
- Zweng M.M., Reagan J.R., Seidov D., Boyer T.P., Locarnini R.A., Garcia H.E., Mishonov A.V., Baranova O.K., Weathers K.W., Paver C.R. & Smolyar I.V. 2019. World Ocean Atlas 2018. Vol. 2. Salinity. NOAA Atlas NESDIS 82. A. Mishonov, technical ed. Silver Spring, MD: National Oceanic and Atmospheric Administration, US Dept. of Commerce.
Related Articles
Agnieszka Promińska
Waldemar Walczowski
Eva Falck