This article is published under a Creative Commons license and not by the author of the article. So if you find any inaccuracies, you can correct them by updating the article.

Aggregations of foraging black guillemots (Cepphus grylle) at a sea-ice edge in front of a tidewater glacier



Øystein Varpe
Geir W. Gabrielsen
Published: Nov. 9, 2022
Latest article update: July 26, 2023
This article is published under the license




Abstract
Seabirds in cold biomes sometimes aggregate near glacier fronts and at sea-ice edges to forage. In this note, we report on large aggregations of black guillemots (Cepphus grylle) at the edge of sea ice in front of the tidewater glacier Kongsbreen (Kongsfjorden, Svalbard). During several days in the second half of June 2011, we observed 49–155 individuals of black guillemots at this ice edge. They foraged actively, and many of the dives were directed underneath the sea ice. The outflow of glacial meltwater and resulting upwelling generated opportunities for the black guillemots to feed, likely on zooplankton or fish. The black guillemots used the sea ice as a resting platform between dives or diving sessions, and whilst on the ice, they interacted socially. On our last visit, the sea ice was gone, and the black guillemots had left the bay. At the neighbouring tidewater glacier Kronebreen, there was no sea ice connected to the glacier. Surface-feeding seabirds, particularly black-legged kittiwakes (Rissa tridactyla), were numerous at the plumes generated by meltwater from Kronebreen. Black guillemots were not seen at these plumes, but some individuals were seen scattered in the fjord system. Our observations add to the natural history of black guillemots and enhance our knowledge of ecological interactions and seabird habitat use shaped by tidewater glaciers.
Keywords
Arctic, marine-terminating glacier, Alcidae, cryosphere, coastal ecology, estuarine circulation
Introduction
The distribution of animals in space and time is the outcome of complex interactions between external states, such as food availability, predation risk and abiotic conditions, and internal states, such as hunger, body condition and breeding status (Levin 1992; Houston & McNamara 1999). For foraging animals, aggregations are likely where food is abundant and available (Fretwell & Lucas 1970; Stephens et al. 2007). Seabirds are known to aggregate in the Arctic marginal ice zone because of the good feeding conditions (Bradstreet 1979, 1980; Lønne & Gabrielsen 1992; Divoky et al. 2016), with similar patterns in the Antarctic (e.g., Tarroux et al. 2020). Arctic seabirds, particularly surface feeders, also aggregate near tidewater glaciers (also known as marine-terminating glaciers) in the melting season (McLaren & Renaud 1982; Day et al. 2000; Lydersen et al. 2014; Stempniewicz et al. 2017; Nishizawa et al. 2020; Bertrand et al. 2021). Freshwater run-off from tidewater glaciers leads to upwelling and the entrapment of zooplankton, which results in predictable aggregations of prey (Węsławski et al. 2000; Lydersen et al. 2014; Urbanski et al. 2017).
The black guillemot is primarily a coastal and fjord species, diving for food in relatively shallow areas near the coastline, usually solitarily or in small groups (Anker-Nilssen et al. 2000; authors’ own observations). It eats small fish and larger zooplankton, as well as benthic organisms (Lydersen et al. 1989; Lønne & Gabrielsen 1992; Mehlum & Gabrielsen 1993; Divoky et al. 2021). The black guillemot can also be common offshore at sea-ice edges (Bradstreet 1979; Divoky et al. 2016). When found in glacial bays, black guillemots are less closely associated with glacier fronts than surface-feeding species, such as the black-legged kittiwake (Rissa tridactyla; Lydersen et al. 2014; Stempniewicz et al. 2017). In this study, we report on large aggregations of foraging and resting black guillemots in a habitat that combines a well-defined sea-ice edge with meltwater run-off from a tidewater glacier.
Study site and methods
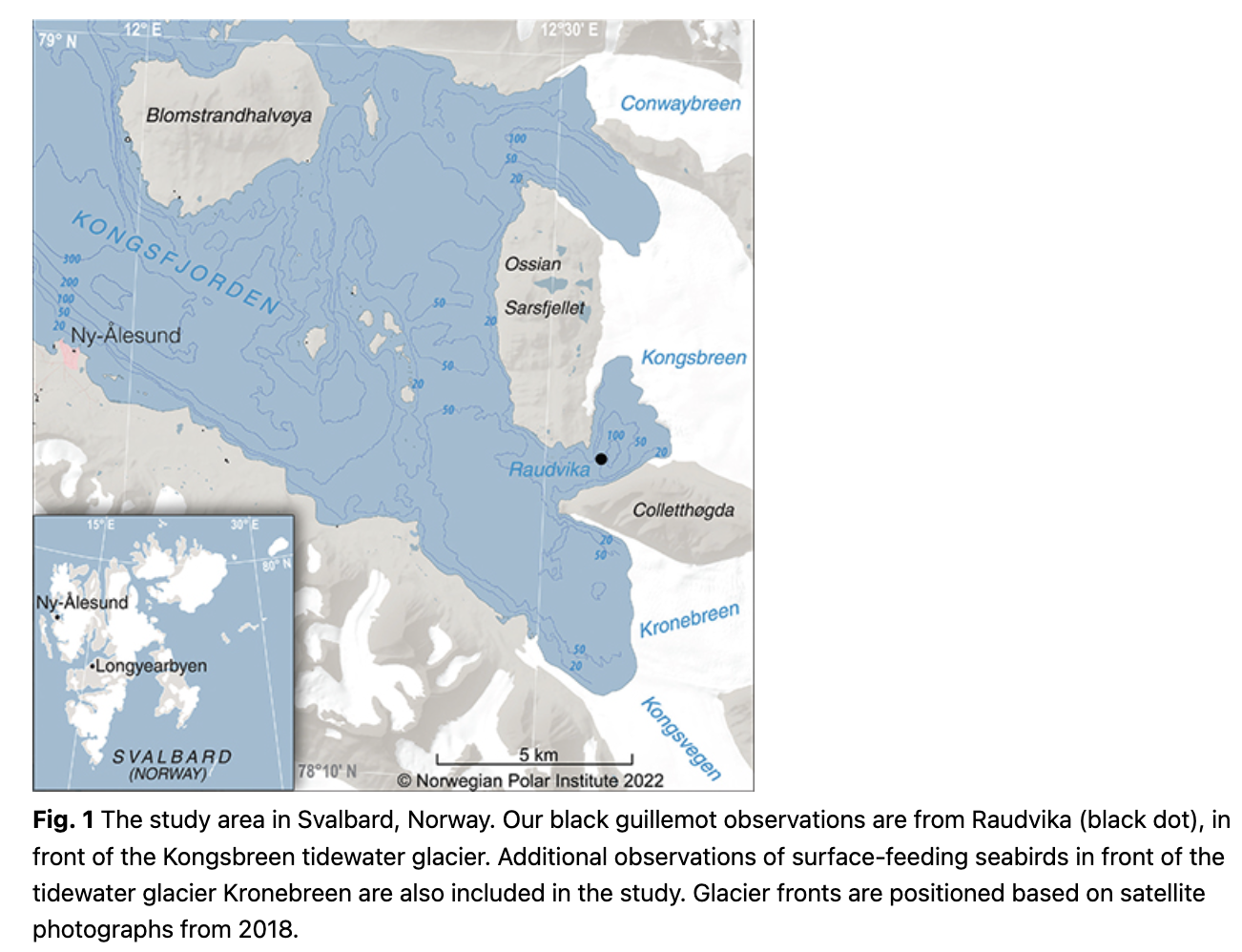
The observations are from Raudvika (78.9°N, 12.6°E), a glacial bay in inner Kongsfjorden, Svalbard, near the research community of Ny-Ålesund (Fig. 1). The tidewater glacier Kongsbreen terminates in this bay. The bay was visited in the second half of June 2011, and the black guillemots counted on four dates (19, 22, 23 and 25 June). The visits to Raudvika were made in a small boat and lasted about 15 min each time, except for a longer visit of about an hour on 22 June, when the guillemot behaviour was observed in some detail and was filmed (Supplementary video). On this longer visit, we anchored the boat to the fast sea ice. Our observations at Raudvika were supplemented with observations of surface-feeding seabirds aggregated at the neighbouring tidal glacier Kronebreen (Fig. 1), 2–3 km away from the ice edge in Raudvika, on 23 June.
The sea ice had many surface melt ponds, was connected to the glacier and had a well-defined edge, and there were no loose ice floes on the bay (Fig. 2). The ice edge was about 1.3 km long and situated parallel to, and about 2 km from, the glacier front (Figs. 1, 2). The position of the ice edge was the same on our three first visits. There was a strong outflow of water, likely glacier meltwater, particularly in the middle of the bay. On the sea ice nearer to the glacier, ringed seals (Pusa hispida) hauled out. On our final visit (25 June), there was no sea ice in the bay.

Observations
The black guillemots were found on the sea ice or on the water near the ice edge (Fig. 2, Supplementary video), mainly at the middle and northern parts of the bay. This distribution was consistent for all three days with sea ice. The number of guillemots on these days ranged 49–155 individuals (Table 1). Only a few scattered black guillemots were observed on the last visit (25 June), after the sea ice had disappeared from Raudvika. We observed a mix of immature and adult individuals (Supplementary Fig. S1), which can be distinguished on the basis of the barred white wing patch of immature individuals (Cramp 1986).
Table 1 Number of black guillemots at the sea-ice edge in Raudvika, Kongsfjorden, on different days during the second half of June 2011. | |||
Date | Number of black guillemots | Comments | Sea-ice conditions |
19 June | Ca. 100 | Approximate count | Well-defined ice edge |
22 June | 155 | We spent about one hour in the bay, and the number of birds were similar during this period (the Supplementary video is from this occasion) | Well-defined ice edge |
23 June | 49 | Zooplankton specimens from surface waters picked nearer to the Kronebreen glacier | Well-defined ice edge |
25 June | 6 | For the whole of Raudvika | No sea ice |
The birds were diving frequently, and some dives were directed underneath the ice. Other dives took place in the ice-free waters just off the ice edge. On several occasions, individuals used the sea ice as a starting point for dives, launching themselves onto the water and after a few seconds initiating the dive. The sea ice also served as a resting platform between dives or diving sessions. A diverse range of social interactions could be observed, such as some birds chasing each other (Supplementary video).
At the neighbouring glacier, Kronebreen, black-legged kittiwakes were very abundant, in addition to some northern fulmars (Fulmarus glacialis), glaucous gulls (Larus hyperboreus) and Arctic terns (Sterna paradisaea). These aggregations are commonly seen at Kronebreen during summer (authors’ observations). Judging from the three main aggregations of birds, meltwater and plume formation occurred at three places along the glacier front (see Supplementary Fig. S2 for one of these aggregations). Because of the risk of ice calving from the glacier, we did not enter these plumes, which are located close (10–100 m) to the glacier front. On the fjord in front of Kronebreen only a few, scattered black guillemots were seen, and they were not associated with the plumes. Some distance from the Kronebreen plumes, we observed macrozooplankton that was slow moving and not behaving normally and could be picked by hand from near the surface. They were probably suffering from low salinity and osmotic stress. We did not perform quantitative sampling, but we picked up a few specimens, which were the amphipod Themisto libellula and krill Thysanoessa sp. (Supplementary Fig. S3).
Discussion
Our observations illustrate how sea ice, with a well-defined ice edge, in front of a melting tidewater glacier sets up favourable feeding conditions for a diving bird. The black guillemots were not evenly distributed along the ice edge but were aggregated near the well-defined outflow that was likely meltwater from the glacier. Plumes and upwelling in front of tidewater glacier fronts are well known to attract large numbers of foraging seabirds (Hartley & Fisher 1936; Stott 1936; Mehlum 1984; Węsławski & Legeżyńska 1998; Węsławski et al. 2000; Lydersen et al. 2014; Bertrand et al. 2021).
Themisto libellula and Thysanoessa sp., which we observed nearer to Kronebreen, have been reported as abundant near glaciers in Kongsfjorden (Węsławski et al. 2000), and their presence near the surface led Węsławski et al. (2000) to suggest that the zooplankton was entrapped by the estuarine circulation, and that the freshwater possibly impairs the zooplankton, making them relatively easy prey (Lydersen et al. 2014). Amphipods, including T. libellula, and other invertebrates are frequently observed as part of the black guillemot diet (Mehlum & Gabrielsen 1993), more so in coastal areas compared to the fish-dominated diet observed offshore in the marginal ice zone (Mehlum & Gabrielsen 1993). Previous studies of seabird aggregations in front of glacier fronts lacked the fast sea ice in front of the glacier that we observed in Raudvika. What we observed in this bay is, however, somewhat similar to aggregations of black guillemots observed further east in Svalbard, on a cruise with the RV Lance in July 2007: aggregations of 100–150 black guillemots were seen feeding in Wallenbergfjorden (Palanderbukta), close to sea ice connected to the glacier Vegafonna (pers. obs. G.W. Gabrielsen).
Black guillemots were the only bird species in the bay with fjord ice, whereas several surface feeding species were present at the plumes by the neighbouring Kronebreen. In a detailed study of seabird distributions in front of a glacier in Greenland, Nishizawa et al. (2020) also report an absence of diving birds, and a large number of surface feeders, at the glacier front plumes. One explanation for the black guillemots’ attraction to the bay with fast ice could be that the food may have primarily been present under the ice or at some depth, making it available only to a diving bird and inaccessible for surface feeders, such as gulls. The prey community may also have differed, potentially with polar cod (Boreogadus saida) being present in Raudvika. This fish species is well known to Kongsfjorden and other fjords in Svalbard (e.g., Nahrgang et al. 2014). Polar cod can be found under sea ice and would benefit from invertebrates in the water current (Crawford & Jorgenson 1993; Hop & Gjøsaeter 2013). Furthermore, the absence of black guillemots at the Kroneebreen plumes may be related to their relative inability to escape quickly when the glacier is calving, in contrast to swift flyers and non-divers such as black-legged kittiwakes.
The waters where the black guillemots foraged were rather turbid (Fig. 2, Supplementary material), raising questions about their vision whilst diving. Water clarity strongly impacts the detection distance of visually searching predators (Aksnes & Utne 1997), and snow-covered sea ice limits the light entering the water column, with many implications for ecological interactions (Varpe et al. 2015; Langbehn & Varpe 2017). The distribution and feeding behaviour of Kittlitz’s murrelets (Brachyramphus brevirostris) in Alaskan fjords suggest that this seabird species is particularly adapted to low-light feeding conditions (Day et al. 2000; Arimitsu et al. 2012). They feed on macrozooplankton and small schooling fishes (Day & Nigro 2000) and also breed close to glaciers. Our observations suggest that the black guillemot, similarly to the murrelet, may have a particular capacity to detect prey under turbid and low-light conditions. Black guillemots with food in their stomachs can be found in Svalbard during the polar night, supporting the idea that they can detect and catch prey under low-light conditions (Berge et al. 2015).
The sea-ice edge was a much-used roosting or resting platform. Diving in cold water and digesting cold prey are energy demanding, and digestion takes time (Hawkins et al. 1997). Resting on sea ice can then be beneficial and makes it easier to regain body temperature after repeated dives in the cold water (Gabrielsen et al. 1988; de Vries & van Eerden 1995; Niizuma et al. 2007). Near the neighbouring Kronebreen, pieces of glacier ice floating in the fjord were much used as resting platforms by black-legged kittiwakes (Supplementary Fig. S4) but were not used by black guillemots, indicating that they had better conditions at the fast ice in Raudvika.
The black guillemots interacted actively when on the sea ice (Supplementary video), and mature and immature individuals were there together. Black guillemots breed in the area, and colonies are situated on Midtholmen and Juttaholmen, some 5–10 km west of Raudvika, with about 20 and 30 breeding pairs, respectively (pers. obs. G.W. Gabrielsen). At the time of our observations (late June), breeding black guillemots would be incubating their two eggs, with hatching around 10–15 July (pers. obs. G.W. Gabrielsen; Mehlum et al. 1993). Some of the black guillemots we observed in Raudvika may have been in between incubation spells.
As global warming impacts the cryosphere, sea ice and glaciers are diminishing in Svalbard and the Barents Sea (Onarheim et al. 2018; Duarte et al. 2020; Geyman et al. 2022). These changes to the ice environment will impact black guillemots and similar birds in diverse ways. Sea-ice extent in Kongsfjorden is variable; in recent years, sea ice has covered only the inner parts of the fjord (Pavlova et al. 2019). If this trend continues, sea-ice edges near the glacier are unlikely to coincide in time with the seasonal meltwater run-off from the glacier. Furthermore, changing sea-ice conditions offshore will impact black guillemots feeding in the marginal ice zone. Loss of sea ice nearshore is related to reduced breeding success and population size of black guillemots in Arctic Alaska (Divoky et al. 2015), along with changing foraging behaviour and prey composition (Divoky et al. 2021). Retreating glaciers have a range of ecological consequences in terrestrial, freshwater and marine environments (Cauvy-Fraunié & Dangles 2019). As they retreat, tidewater glaciers eventually become land-terminating glaciers (Lydersen et al. 2014). These changes will have many implications as a diverse range of ecological interactions occur in front of tidewater glaciers, particularly during the part of the year when freshwater run-off from the glaciers affect the estuarine circulation (Lydersen et al. 2014). Our observations add to the range of such glacier-related ecological interactions by documenting how the fast sea ice (with a well-defined ice edge) in front of a melting tidewater glacier sets up favourable feeding conditions for a diving seabird.
Acknowledgements
The authors thank Stephen J. Coulson for the aerial photograph in Fig. 2c. The authors also thank George Divoky and one anonymous reviewer for valuable comments on a previous version of the manuscript.
References
Aksnes D.L. & Utne A.C.W. 1997. A revised model of visual range in fish. Sarsia 82, 137–147, doi: 10.1080/00364827.1997.10413647. |
Anker-Nilssen T., Bakken V., Strøm H., Golovkin A.N., Bianki V.V. & Tatarinkova I.P. 2000. The status of marine birds breeding in the Barents Sea area. Norwegian Polar Institute Report Series 113. Tromsø: Norwegian Polar Institute. |
Arimitsu M.L., Piatt J.F., Madison E.N., Conaway J.S. & Hillgruber N. 2012. Oceanographic gradients and seabird prey community dynamics in glacial fjords. Fisheries Oceanography 21, 148–169, doi: 10.1111/j.1365-2419.2012.00616.x. |
Berge J., Daase M., Renaud P.E., Ambrose W.G., Jr., Darnis G., Last K.S., Leu E., Cohen J.H., Johnsen G., Moline M.A., Cottier F., Varpe Ø., Shunatova N., Bałazy P., Morata N., Massabuau J.-C., Falk-Petersen S., Kosobokova K., Hoppe C.J.M., Węsławski J.M., Kukliński P., Legeżyńska J., Nikishina D., Cusa M., Kędra M., Włodarska-Kowalczuk M., Vogedes D., Camus L., Tran D., Michaud E., Gabrielsen T.M., Granovitch A., Gonchar A., Krapp R. & Callesen T.A. 2015. Unexpected levels of biological activity during the polar night offer new perspectives on a warming Arctic. Current Biology 25, 2555–2561, doi: 10.1016/j.cub.2015.08.024. |
Bertrand P., Bêty J., Yoccoz N.G., Fortin M.-J., Strøm H., Steen H., Kohler J., Harris S.M., Patrick S.C., Chastel O., Blévin P., Hop H., Moholdt G., Maton J. & Descamps S. 2021. Fine-scale spatial segregation in a pelagic seabird driven by differential use of tidewater glacier fronts. Scientific Reports 11, article no. 22109, doi: 10.1038/s41598-021-01404-1. |
Bradstreet M.S.W. 1979. Thick-billed murres and black guillemots in the Barrow Strait area, N.W.T., during spring: distribution and habitat use. Canadian Journal of Zoology 57, 1789–1802, doi: 10.1139/z79-233. |
Bradstreet M.S.W. 1980. Thick-billed murres and black guillemots in the Barrow Strait area, NWT, during spring: diets and food availability along ice edges. Canadian Journal of Zoology 58, 2120–2140, doi: 10.1139/z80-292. |
Cauvy-Fraunié S. & Dangles O. 2019. A global synthesis of biodiversity responses to glacier retreat. Nature Ecology & Evolution 3, 1675–1685, doi: 10.1038/s41559-019-1042-8. |
Cramp S. (ed.) 1986. Handbook of the birds of Europe the Middle Ease and North Africa: the birds of the Western Palearctic. Volume IV: terns to woodpeckers. Oxford: Oxford University Press. |
Crawford R.E. & Jorgenson J.K. 1993. Schooling behaviour of Arctic cod, Boreogadus saida, in relation to drifting pack ice. Environmental Biology of Fishes 36, 345–357, doi: 10.1007/BF00012412. |
Day R.H. & Nigro D.A. 2000. Feeding ecology of Kittlitz’s and marbled murrelets in Prince William Sound, Alaska. Waterbirds 23, 1–14. |
Day R.H., Nigro D.A. & Prichard A.K. 2000. At-sea habitat use by the Kittlitz’s murrelet Brachyramphus brevirostris in nearshore waters of Prince William Sound, Alaska. Marine Ornithology 28, 105–114. |
de Vries J.D. & van Eerden M.R. 1995. Thermal conductance in aquatic birds in relation to the degree of water contact, body mass, and body fat: energetic implications of living in a strong cooling environment. Physiological Zoology 68, 1143–1163, doi: 10.1086/physzool.68.6.30163797. |
Divoky G.J., Brown E. & Elliott K.H. 2021. Reduced seasonal sea ice and increased sea surface temperature change prey and foraging behaviour in an ice-obligate Arctic seabird, Mandt’s black guillemot (Cepphus -grylle mandtii). Polar Biology 44, 701–715, doi: 10.1007/s00300-021-02826-3. |
Divoky G.J., Douglas D.C. & Stenhouse I.J. 2016. Arctic sea ice a major determinant in Mandt’s black guillemot movement and distribution during non-breeding season. Biology Letters 12, article no. 20160275, doi: 10.1098/rsbl.2016.0275. |
Divoky G.J., Lukacs P.M. & Druckenmiller M.L. 2015. Effects of recent decreases in Arctic sea ice on an ice-associated marine bird. Progress in Oceanography 136, 151–161, doi: 10.1016/j.pocean.2015.05.010. |
Duarte P., Sundfjord A., Meyer A., Hudson S.R., Spreen G. & Smedsrud L.H. 2020. Warm Atlantic Water explains observed sea ice melt rates north of Svalbard. Journal of Geophysical Research—Oceans 125, e2019JC015662, doi: 10.1029/2019JC015662. |
Fretwell S.D. & Lucas H.L., Jr. 1970. On territorial behavior and other factors influencing habitat distribution in birds: I. Theoretical development. Acta Biotheoretica 19, 16–36, doi: 10.1007/BF01601953. |
Gabrielsen G.W., Mehlum F. & Karlsen H.E. 1988. Thermoregulation in four species of Arctic seabirds. Journal of Comparative Physiology B 157, 703–708, doi: 10.1007/BF00691000. |
Geyman E.C., van Pelt W.J.J., Maloof A.C., Aas H.F. & Kohler J. 2022. Historical glacier change on Svalbard predicts doubling of mass loss by 2100. Nature 601, 374–379, doi: 10.1038/s41586-021-04314-4. |
Hartley C.H. & Fisher J. 1936. The marine foods of birds in an inland fjord region in west Spitsbergen: part 2. Birds. Journal of Animal Ecology 5, 370–389, doi: 10.2307/1041. |
Hawkins P.A.J., Butler P.J., Woakes A.J. & Gabrielsen G.W. 1997. Heat increment of feeding in Brunnich’s guillemot Uria lomvia. Journal of Experimental Biology 200, 1757–1763, doi: 10.1242/jeb.200.12.1757. |
Hop H. & Gjøsaeter H. 2013. Polar cod (Boreogadus saida) and capelin (Mallotus villosus) as key species in marine food webs of the Arctic and the Barents Sea. Marine Biology Research 9, 878–894, doi: 10.1080/17451000.2013.775458. |
Houston A.I. & McNamara J.M. 1999. Models of adaptive behaviour. Cambridge: Cambridge University Press. |
Langbehn T.J. & Varpe Ø. 2017. Sea-ice loss boosts visual search: fish foraging and changing pelagic interactions in polar oceans. Global Change Biology 23, 5318–5330, doi: 10.1111/gcb.13797. |
Levin S.A. 1992. The problem of pattern and scale in ecology. Ecology 73, 1943–1967, doi: 10.2307/1941447. |
Lønne O.J. & Gabrielsen G.W. 1992. Summer diet of seabirds feeding in sea-ice covered waters near Svalbard. Polar Biology 12, 685–692, doi: 10.1007/BF00238868. |
Lydersen C., Assmy P., Falk-Petersen S., Kohler J., Kovacs K.M., Reigstad M., Steen H., Strøm H., Sundfjord A., Varpe Ø., Walczowski W., Weslawski J.M. & Zajaczkowski M. 2014. The importance of tidewater glaciers for marine mammals and seabirds in Svalbard, Norway. Journal of Marine Systems 129, 452–471, doi: 10.1016/j.jmarsys.2013.09.006. |
Lydersen C., Gjertz I. & Węsławski J.M. 1989. Stomach contents of autumn-feeding marine vertebrates from Hornsund, Svalbard. Polar Record 25, 107–114, doi: 10.1017/S0032247400010408. |
McLaren P.L. & Renaud W.E. 1982. Seabird concentrations in late summer along the coasts of Devon and Ellesmere Island, NWT. Arctic 35, 112–117, doi: 10.14430/arctic2311. |
Mehlum F. 1984. Konsentrasjoner av sjøfugl langs kanten av isbreer og utenfor breelver på Svalbard. (Concentrations of seabirds along the edges of glaciers and off glacial streams in Svalbard.) Fauna 37, 156–160. |
Mehlum F. & Gabrielsen G.W. 1993. The diet of High‐Arctic seabirds in coastal and ice‐covered, pelagic areas near the Svalbard archipelago. Polar Research 12, 1–20, doi: 10.1111/j.1751-8369.1993.tb00417.x. |
Mehlum F., Gabrielsen G.W. & Nagy K.A. 1993. Energy expenditure by black guillemots (Cepphus grylle) during chick-rearing. Colonial Waterbirds 16, 45-52, doi: 10.2307/1521555. |
Nahrgang J., Varpe Ø., Korshunova E., Murzina S., Hallanger I.G., Vieweg I. & Berge J. 2014. Gender specific reproductive strategies of an Arctic key species (Boreogadus saida) and implications of climate change. PLoS One 9, e98452, doi: 10.1371/journal.pone.0098452. |
Niizuma Y., Gabrielsen G.W., Sato K., Watanuki Y. & Naito Y. 2007. Brunnich’s guillemots (Uria lomvia) maintain high temperature in the body core during dives. Comparative Biochemistry and Physiology, Part A 147, 438–444, doi: 10.1016/j.cbpa.2007.01.014. |
Nishizawa B., Kanna N., Abe Y., Ohashi Y., Sakakibara D., Asaji I., Sugiyama S., Yamaguchi A. & Watanuki Y. 2020. Contrasting assemblages of seabirds in the subglacial meltwater plume and oceanic water of Bowdoin Fjord, northwestern Greenland. ICES Journal of Marine Science 77, 711–720, doi: 10.1093/icesjms/fsz213. |
Onarheim I.H., Eldevik T., Smedsrud L.H. & Stroeve J.C. 2018. Seasonal and regional manifestation of Arctic sea ice loss. Journal of Climate 31, 4917–4932, doi: 10.1175/jcli-d-17-0427.1. |
Pavlova O., Gerland S. & Hop H. 2019. Changes in sea-ice extent and thickness in Kongsfjorden, Svalbard (2003–2016). In H. Hop & C. Wiencke (eds.): The ecosystem of Kongsfjorden, Svalbard. Pp. 105–136. Cham: Springer International Publishing. |
Stempniewicz L., Goc M., Kidawa D., Urbański J., Hadwiczak M. & Zwolicki A. 2017. Marine birds and mammals foraging in the rapidly deglaciating Arctic fjord—numbers, distribution and habitat preferences. Climatic Change 140, 533–548, doi: 10.1007/s10584-016-1853-4. |
Stephens D.W., Brown J.S. & Ydenberg R.C. 2007. Foraging: behaviour and ecology. Chicago, IL: University of Chicago Press. |
Stott F.C. 1936. The marine foods of birds in an inland fjord region in west Spitsbergen: part 1. plankton and in shore benthos. Journal of Animal Ecology 5, 356–369, doi: 10.2307/1040. |
Tarroux A., Cherel Y., Fauchald P., Kato A., Love O.P., Ropert-Coudert Y., Spreen G., Varpe Ø., Weimerskirch H., Yoccoz N.G., Zahn S. & Descamps S. 2020. Foraging tactics in dynamic sea-ice habitats affect individual state in a long-ranging seabird. Functional Ecology 34, 1839–1856, doi: 10.1111/1365-2435.13632. |
Urbanski J.A., Stempniewicz L., Węsławski J.M., Dragańska-Deja K., Wochna A., Goc M. & Iliszko L. 2017. Subglacial discharges create fluctuating foraging hotspots for sea birds in tidewater glacier bays. Scientific Reports 7, article no. 43999, doi: 10.1038/srep43999. |
Varpe Ø., Daase M. & Kristiansen T. 2015. A fish-eye view on the new Arctic lightscape. ICES Journal of Marine Science 72, 2532–2538, doi: 10.1093/icesjms/fsv129. |
Węsławski J.M. & Legeżyńska J. 1998. Glaciers caused zooplankton mortality? Journal of Plankton Research 20, 1233–1240, doi: 10.1093/plankt/20.7.1233 |
Węsławski J.M., Pedersen G., Falk-Petersen S. & Poraziński K. 2000. Entrapment of macroplankton in an Arctic fjord basin, Kongsfjorden, Svalbard. Oceanologia 42, 57–69. |
Related Articles
Shengni Duan
Zhina Jiang
Min Wen
Olof Bengtsson
Kit M. Kovacs
Christian Lydersen
Michał Hubert Węgrzyn
Beata Cykowska-Marzencka
Monica Alterskjær Sundset
Paulina Wietrzyk-Pełka
et al.
Clémentine Peggy Anne-Marie Colpaert
Boris Leonidovich Nikitenko
Heidi Ahonen
Kathleen M. Stafford
Christian Lydersen
Laura de Steur
et al.