This article is published under a Creative Commons license and not by the author of the article. So if you find any inaccuracies, you can correct them by updating the article.

Плата за решение задачи: биофизические предпосылки и возможные эволюционные последствия



Алексей Крушинский
Published: Jan. 31, 2015
Latest article update: Sept. 15, 2022
This article is published under the license




Abstract
40 лет назад Л. В. Крушинским была опубликована дискуссионная статья «О возможном механизме рассудка», в которой он предположил, что упорядоченность мозга и избыточность нейронов должны быть тем выше, чем сложнее когнитивные задачи, решаемые животным. Анализ термодинамической и биофизической литературы свидетельствует о том, что у этой мысли есть определенные физические предпосылки, разбору которых я посвящаю эту статью. Основываясь на положениях Э. Шредингера, Л. Больцмана, Л. Бриллюэна, К. Шеннона и др., можно показать, что за решение логической задачи надо заплатить увеличением энтропии мозга, то есть снижением его упорядоченности, превышающем информацию, содержащуюся в задаче. Поэтому если мозг не обладает некоторым запасом упорядоченности, он просто не сможет решить задачу. Биофизический вывод о том, что мозг обладает запасом упорядоченности, превосходящим информационную сложность решаемых им задач, позволяет сделать предположения о возможной роли этого явления в биологической эволюции нервной системы.
Keywords
Информация, эволюция когнитивных функций, экстраполяция, энтропия, мышление, решение задачи
Памяти Леонида Викторовича Крушинского 1911-1984
Беспорядок неотделим от «творчества», поскольку это последнее характеризуется определенным «порядком».
Поль Валери
Концепция элементарной рассудочной деятельности Л.В. Крушинского является одной из составляющих общей теории поведения. Наиболее полное развитие эта концепция получила в книге «Биологические основы рассудочной деятельности» (Крушинский Л. В., 1977, 1986). Экспериментально доказав наличие элементарного разума у животных, он показал адаптивное значение этой функции мозга и ее роль в процессе эволюции высших позвоночных животных и человека.
Как ученого Леонида Викторовича прежде всего характеризовала глубокая культура и широта научных интересов. Значительное влияние на развитие его научных взглядов оказали идеи крупнейших физиков Э. Шредингера и Н. Бора о возможности применения физических законов к биологическим системам, а также работы выдающегося математика А.А. Ляпунова, с которым его связывала многолетняя дружба. Поздний период творчества Л.В. Крушинского характеризуется стремлением к максимально возможной точности и дискретности в описании тех физиологических механизмов, которые лежат в основе поведения животных. Так, гипотеза о возможных механизмах элементарной рассудочной деятельности (Крушинский Л. В., 1974, 2013; Крушинский А. Л., 2013; Полетаева, 2013) в значительной степени определяет возможность описания некоторых ее положений 1 (В исходной публикации Л.В. Крушинского гипотеза сформулирована в виде пяти положений (Прим. ред.).) с использованием точных соотношений термодинамики и теории информации. Целью моей работы было подробнее развить именно эти аспекты работы Л.В. Крушинского на основе ставших уже классическими положениях Э. Шредингера, Л. Больцмана, Л. Бриллюэна, К. Шеннона и др.
Согласно гипотезе Л.В. Крушинского (1974), в основе разумного поведения лежит возможность решения животными новых для них задач, построенных на простейших понятиях о пространстве и движении. Одним из наиболее существенных звеньев гипотезы является положение о том, что необходимым условием для решения новых задач является избыточное количество нейронов в мозге. Используется общепринятое представление об избыточности как о наличии большего количества нейронов, чем то, которое могло бы обеспечить нормальное функционирование организма. К выводу о наличии избыточности в мозге пришли на основании данных о том, что ряд патологических процессов у человека, сопровождающихся гибелью огромного числа нейронов, может протекать бессимптомно (например, Davie, 2008). «Избыточность» нейронов обнаружена и в простых нервных системах беспозвоночных, например (Zecevic et al., 1989), что не противоречит предположению о связи избыточности с решением новых задач. Теория самоорганизующихся адаптивных систем предполагает, что 1 В исходной публикации Л. В. Крушинского гипотеза сформулирована в виде пяти положений (Прим. ред.). способность снижать неопределенность внешней среды должна быть вообще присуща биологическим организмам (Fristen, 2010).
Сопоставление успеха решения логических задач с нейронной организацией конечного мозга у позвоночных показало, что чем больше размер мозга, то есть количество нейронов, из которых он состоит, и чем сложнее и совершеннее система соединений между ними, тем больше вероятность того, что животное обладает развитой способностью к элементарному мышлению. Экспериментальные данные (Попова и др., 1983; Перепелкина и др., 2006) хорошо согласуются с положением о роли избыточности нейронов для осуществления разумного поведения. Ссылаясь на Э. Шредингера (1947), Л.В. Крушинский предположил, что для восприятия структурной организации среды процессы, происходящие в мозге, должны быть упорядоченными, причем эта упорядоченность тем выше, чем большее число нейронов вовлечено в процесс мышления. В последнее время вопрос о том, что следует понимать под «высокой упорядоченностью мозга» активно дискутируется (Huang, 2008; Fristen, 2010; Carhart-HarrisetaL, 2014). Среди конкретных факторов, которые следует учитывать при оценке сложности и упорядоченности, называются: количество нейронов, межнейрональных связей (близких и далеких), дендритных шипиков, число и эффективность синапсов, разнообразие рецепторов и сигнальных молекул и т.д. (например, Bargmann, 2012; Toga, 2012; Сахаров, 2012).
Оба предположения Л.В. Крушинского: об избыточности нейронов и о том, что для восприятия структурной организации среды процессы, происходящие в мозге, должны быть упорядоченными, — можно рассмотреть более детально с биофизической точки зрения. Ссылки на хорошо известные положения, которые можно найти в учебных пособиях по биофизике, статистической физике и оригинальных исследованиях по термодинамике неравновесных систем (Блюмен- фельд, 1974; Волькенштейн, 1975, 1980, 2008; Васильев, 1980; Рубин, 1987; Хакен, 1991; Пригожин, Стенгере, 2000) не привожу.
Согласно современным представлениям, биологические системы можно определить как открытые2 (Открытые системы — системы, которые обмениваются энергией и массой (веществом) с окружающей средой), далекие от равновесия, устойчивые системы, способные к размножению (Волькенштейн, 1975; Зотин А.И., Зо- тин А.А., 1999). Э. Шредингер в своей книге «Что такое жизнь с точки зрения физики» (1947), впервые вышедшей в 1944 году, достаточно четко показал возможность описания живых систем при помощи таких физических величин, как энергия и энтропия.
В физических изолированных системах3 (Изолированные системы полностью изолированы, т.е. не обмениваются с окружающей средой энергией и массой.) , согласно второму закону термодинамики, энтропия (S) всегда растет, то есть

достигая максимума в состоянии равновесия. Энтропия физической системы описывается уравнением Больцмана:

где W — термодинамическая вероятность, которая характеризует число микросостояний системы, соответствующих данному макросостоянию; к — постоянная Больцмана, к=1.38-10‘16 эрг-град-1 или 3.31-10-24 энтропийных единиц (э. е.). Макросостояние физической системы определяется такими параметрами, как объем, давление, температура, химический состав и т.п. Согласно Э. Шредингеру, W является мерой неупорядоченности системы, обратная величина 1 /W может рассматриваться как мера упорядоченности. Таким образом, уравнение Больцмана можно написать так:

Величина -S есть мера упорядоченности системы.
Одной из основных идей Шредингера является положение о высокой упорядоченности живых организмов. В процессе жизнедеятельности организм непрерывно увеличивает свою энтропию, приближаясь к состоянию максимальной энтропии, которое является смертью. Он может поддерживать высокую упорядоченность (низкую энтропию), используя свободную энергию4 (Свободная энергия (F) — та часть энергии системы, которая может быть использована для совершения работы: F=U–TS, где U —полная энергия, T — абсолютная температура, S — энтропия.) , содержащуюся в пище, и увеличивая энтропию окружающей среды. Согласно Шредингеру, физические взаимодействия между живым организмом и внешней средой должны обладать известной степенью упорядоченности, то есть подчиняться строгим физическим законам. Такое условие может выполняться только в системах, состоящих из большого количества атомов5 (Упорядоченность организма, состоящего из небольшого числа атомов, уничтожалась бы из-за тепловых флуктуаций (Шредингер, 1947; Волькенштейн, 1975).) . Реальные расчеты оценки упорядоченности достаточно сложны и основаны на приблизительных расчетах количества информации, содержащейся в живом организме6 (Приблизительная оценка количества информации, содержащаяся в организме человека, была определена Блюменфельдом (1974), и, согласно его расчетам, приблизительно равна 1.34026 бит или 300 кал/град (300 э.е.). На основе этой оценки Блюменфельд заключает, что возникновение и усложнение биологической организации происходит практически «бесплатно» и с легкостью компенсируется тривиальными физическими и химическими процессами. По мнению других ученых, такая оценка значительно занижена (Волькенштейн, 2008). Расчеты Блюменфельда дают достаточно логичную оценку структурной информации, необходимой для построения условного организма, состоящего из соизмеримого с человеческим организмом количества клеток. Однако даже если говорить о структурной информации, эта оценка значительно занижена, поскольку не учитывает информацию, содержащуюся в тонкой организации мозга человека. Кроме того, расчеты Блюменфельда не учитывают той информации, которая необходима для описания сложных процессов метаболизма, которые реализуются в живом организме. Таким образом, точная количественная оценка упорядоченности биологической системы достаточно сложна и неоднозначна. Однако высказанное Шредингером положение о высокой упорядоченности мозга, по-видимому, справедливо.).
Работа Шредингера оказала значительное влияние на развитие современной биофизики и термодинамики неравновесных систем. Следует отметить, что, хотя в целом положение Шредингера о высокой упорядоченности живых организмов разделяется многими учеными (Волькенштейн, 1975, 2008; Зотин А.И., Зотин А.А., 1999; Тринчер, 1965; Хастлер, 1967; Шмальгаузен, 1968; Ленинджер, 1985; Грин, 2000), высказываются мнения о том, что применение энтропии для описания этой упорядоченности имеет значительные ограничения (Волькенштейн, 1980; Чернявский, 2004). С другой стороны, понятие энтропии может быть применено к биологическим системам; так, энтропия входит в уравнение для изменения количества тепла, образующегося при диссипации энергии в метаболических процессах (Зотин А.И., Зотин А.А., 1999; Тринчер, 1965; Эбелинг и др., 2001). Дальнейшее развитие идеи Шредингера получили в результате синтеза статистической физики, термодинамики и теории информации.
У истоков этого направления, прежде всего, лежат работы выдающегося физика Л. Бриллюэна (1951,1960, 1966). Ему удалось связать выдвинутое Шредингером понятие упорядоченности (отрицательной энтропии) с информацией. Так, вместо отрицательной энтропии он предложил использовать термин негэнтропия (N) как меру упорядоченности системы:

Бриллюэн показал, что негэнтропия может переходить в информацию и обратно. Это положение лежит в основе сформулированного им негэнтропийного принципа информации, согласно которому информация вносит отрицательный вклад в энтропию. Этот принцип, необходимый для понимания процессов, лежащих в основе получения информации, требует более подробного изложения.
Основные положения этого принципа будут рассмотрены мною на основе работ Л. Бриллюэна (1966), Л.А. Блюменфельда (1974), М.В. Волькенштейна (1975, 1980), В. Эбелинга, А. Энгеля, Р. Файстеля (2001). Информацию можно определить как меру количества неопределенности, которая уничтожается после получения сообщения. Количество информации, содержащейся в любом сообщении, равно

где р0 — априорная вероятность некоторого события до получения сообщения, a p1 — после получения сообщения. Если сообщение достоверно, то есть p1 = 1 то

В качестве единицы информации (бит) принимается количество информации, содержащееся в достоверном сообщении о событии, априорная вероятность которого равна 14. Для выражения информации при описании физических систем используется натуральный логарифм In (логарифм по основанию е). Целесообразность введения логарифма в формулы, описывающие информацию и энтропию, определяется тем, что это придает данным величинам свойство аддитивности (Бриллюэн, 1960,1966).
Энтропия системы, определяемая по уравнению Больцмана (2), связана с информацией следующим образом. Пусть число микросостояний, которые реализуют физическую систему, равно W, а вероятность каждого микросостояния р0=1/W. Допустим, что мы экспериментально, путем множества измерений точно установили, в каком микросостоянии находится система, то есть вероятность ее состояния стала pt=1 (в реальности такой эксперимент невозможен). Полученная информация, согласно (5) равна
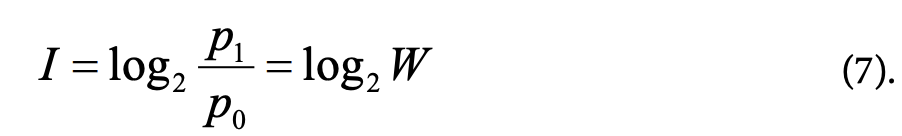
Формулы (2) и (7) совпадают с точностью до постоянного множителя.
Из этого следует, что энтропия системы эквивалентна количеству информации, недостающему для ее полного описания7 (Информация связана с энтропией соотношением I+S=const. Чтобы перейти от информации (в битах) к энтропии (в энтропийных единицах) надо перейти от логарифма при основании 2 к натуральному логарифму и умножить его на k: S=1/log2e·kI (Блюменфельд, 1974).) .
Перенос информации между системами (частями системы) всегда связан с переносом энергии и энтропии. Исследования Л. Бриллюэна показали, что количество переданной информации связано с уменьшением неопределенности системы (Н) соотношением:

При этом неопределенность для системы с п состояниями задается формулой, предложенной А. Шенноном:
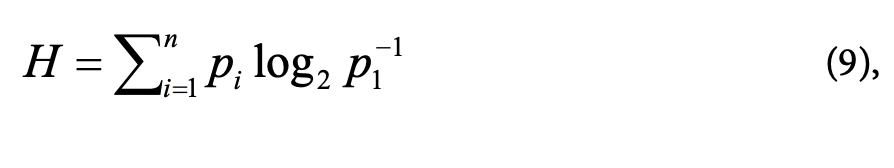
где pi — вероятность нахождения системы в одном из возможных состояний.
Изменение неопределенности связано с изменением энтропии формулой
![]()
Величина Н вычисляется с помощью натуральных логарифмов.
Следовательно, информация эквивалентна отрицательной энтропии (негэнтропии).
Согласно негэнтропийному принципу, всякая информация, извлеченная из эксперимента (наблюдения), должна быть оплачена энтропией. «Если эксперимент дает информацию AJ, то в аппаратуре или в лаборатории, где проводится эксперимент, энтропия должна возрасти» (Бриллюэн, 1966/2006, с. 62), то есть
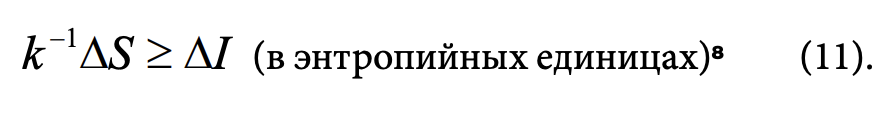
Поскольку, согласно (4), прирост энтропии AS означает убыль общей негэнтропии, информация оплачивается тем, что количество негэнтропии становится меньше. Соотношение (11) также показывает стоимость информации в термодинамических единицах. Один бит информации равен kln2 = 2.3-10-24 э.е., то есть очень малой термодинамической величине. Эта величина соответствует минимальной стоимости одного бита информации. Минимальное количество энергии, требуемое на получение одного бита, определяется по формуле8 (По мнению М.В. Волькенштейна (2008), биологические системы в процессе эволюции приближаются к состоянию, в котором они «платят» по kTln2 за один бит ценной информации и не «платят» ничего за невоспринимаемую информацию, не обладающую ценностью. В принципе разделяя эту точку зрения, добавлю, что вопрос о ценности информации достаточно сложен и неоднозначен.) :
![]()
где Т — абсолютная температура.
Неравенство (11), очевидно, позволяет оценить ту минимальную цену (возрастание энтропии), которой мозг расплачивается за приобретение, переработку и создание новой информации — процессы, которые сопровождают различные формы поведения. Однако вопрос оценки реального возрастания энтропии и затрат энергии при получении информации достаточно сложен. Так, по мнению ряда авторов (Романовский и др., 1984), эта величина может в десятки раз превышать минимальное значение, то есть в реальных системах количество энтропии, возрастающей при получении (запоминании) информации, значительно больше минимального, таким образом
£-1Д5 » AZ (13).
Вернемся к негэнтропийному принципу Бриллюэна, согласно которому любое получение информации оплачивается повышением энтропии в другой части системы. Невозможно получить информацию об изолированной системе. Поток информации всегда сопровождается пропорциональным ему потоком энтропии. Согласно второму закону термодинамики, энтропия в реальных изолированных системах может только возрастать, а информация — только утрачиваться. Информация может приобретаться только в системах, отдающих энтропию во внешнюю среду, то есть для этих систем необходимы неравновесные условия (Бриллюэн, 1966; Эбелинг и др., 2001). Биологические системы являются открытыми, они способны получать, хранить, перерабатывать и производить новую информацию9 (Основываясь на работах В.Эбелинга, М.В. Волькенштейна, Д.С. Чернавского и Ю.М. Романовского, опишем основные свойства, которым должны удовлетворять информационные системы (в том числе и биологические) (Волькенштейн, 2008; Чернавский, 2004; Эбелинг и др., 2001). В таких системах существует много дискретных стационарных состояний (аттракторов). Информационная емкость системы, измеренная в битах, определяется по формуле I=log2n, где n — число состояний. Устойчивость состояний реализуется диссипативными механизмами и практически исключает обусловленный флуктуациями переход. Переход системы из одного стационарного состояния в другое осуществляется в результате внешнего воздействия и лежит в основе механизмов памяти. Одним из необходимых условий осуществления этих механизмов является область, из которой достижимы все состояния (аттракторы) системы. Достижимость определенного аттрактора (группы аттракторов) определяется внешними воздействиями и флуктуациями внутри системы.).
Рассмотрим те соотношения между информацией и энтропией, которые, вероятно, имеют место при разумном поведении животных. Так, согласно гипотезе Л. В. Крушинского, в основе рассудочной деятельности животных лежит их способность улавливать простейшие эмпирические законы, связывающие предметы и явления внешней среды. Реализация разумного поведения, по-видимому, состоит из следующих основных составляющих: 1 — получение и запоминание необходимой и достаточной для решения логической задачи информации ДІр Этот процесс оплачивается возрастанием энтропии в мозге jf'ASj » А/,; 2 — на основе полученной информации (AIJ происходит создание новой информации (АІ2), лежащей в основе алгоритма, реализующего решение данной задачи. Другими словами, АІ2 — это информация о законах, связывающих предметы и явления внешней среды. В результате энтропия мозга возрастает на величину AS2. Реально, согласно (13), &-1Д52 » А/2. Таким образом, общее увеличение энтропии будет описываться неравенством:
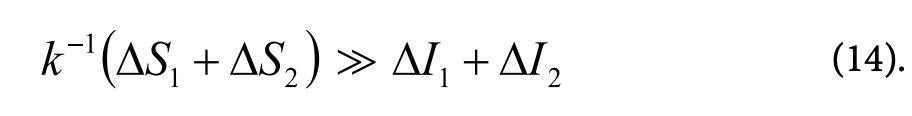
Очевидно, количественные изменения энтропии и информации, происходящие в результате первой стадии, очень малы. Вторая стадия, по-видимому, описывает собственно процесс мышления. Думаю, что количество информации, создающееся в результате решения логической задачи (АІ2), и соответствующий прирост энтропии зависит от сложности задачи и может измеряться значительной величиной. В результате предшествующего опыта животное (или человек) может располагать некоторой информацией (Іо), которая составляет часть информации, содержащейся в решаемой задаче. Очевидно, такая информация может облегчить решение задачи. Соответственно, энтропийная плата за решение задачи уменьшится на величину А^о Ä к^о-
Основной вывод из описанных выше рассуждений, по-видимому, можно сформулировать следующим образом: за решение логической задачи надо заплатить увеличением энтропии мозга, которое выражается неравенством (13), то есть k~lAS » AI (S » LAZ).
Высокая упорядоченность мозга и его функций, возникшая на основе избыточного количества нейронов, по мнению Л.В. Крушинского, является необходимым условием для осуществления мышления. Возрастание энтропии мозга при решении логической задачи, согласно неравенству (13), по существу означает понижение его упорядоченности (негэнтропии). Поэтому, если мозг не обладает некоторым запасом упорядоченности, он просто не сможет заплатить по цене AS » кАІ за создание той информации, которая лежит в основе логической задачи (физического закона), и задача в принципе не может быть решена. Так, рыбы в принципе не способны к решению экстраполяционной задачи без предварительного обучения. С этой задачей успешно справляются животные с более совершенным строением мозга (некоторые рептилии, хищные млекопитающие, птицы из семейства вороновых). Более сложную задачу, связанную с оперированием геометрической размерностью фигур, могут решать только животные, обладающие более развитым мозгом. В целом пределы проявления мышления значительно шире у животных, имеющих более сложный мозг (Крушинский, 1977; Зорина, 2005).
Часть свободной энергии, затраченной на решение логической задачи, необратимо переходит в тепловую энергию. Это количество тепла определяется по формуле AQ = TAS. Реально, учитывая (13), получим АQ » кТАІ. Поскольку живой организм представляет собой открытую систему, повышение энтропии мозга в результате решения задачи в дальнейшем может компенсироваться за счет свободной энергии, содержащейся в пище, и отвода образовавшегося тепла в окружающую среду. Может ли при этом нервная система перейти на более высокий уровень упорядоченности? По-видимому, да. Однако хочу подчеркнуть, что однозначно определить, как изменится упорядоченность мозга в целом после успешного решения задачи и компенсаторного повышения энтропии внешней среды сложно. Она может возрасти, не измениться или даже снизиться, несмотря на то что была приобретена информация (за счет снижения упорядоченности в других отделах мозга). Наиболее уверенно можно говорить о том, что следствием решения задачи будет перераспределение упорядоченности в мозге.
Несмотря на низкую стоимость информационных процессов в единицах энтропии и энергии, не исключено, что возрастание энтропии и диссипация энергии, сопровождающие мышление, могут нарушать нормальную работу мозга и приводить к невротическим срывам. Увеличение энтропии внешней среды не является мгновенным и очевидным событием. В реальности имеет место некоторая многоступенчатая последовательность событий, приводящая к росту энтропии внешней среды и компенсации энергетической стоимости. По Бриллюэну, «поток информации всегда сопровождается пропорциональным ему потоком энтропии» (Бриллюэн, 1966). Очевидно, что начинаться этот поток будет максимально близко к непосредственному акцептору информации, то есть с подсистем ЦНС, возможно весьма локальных, имеющих непосредственное отношение к получению данной информации. Компенсировать за счет энергии утрату упорядоченности в другом отделе ЦНС, по-видимому, не всегда удается. Получение «сверхновой» или сверхсложной информации может оплачиваться безвозвратной утратой некоторой исходной информации, необходимой для обеспечения базовых функций. Энергетическая плата за любую операцию, в том числе решение задачи и получение новой информации, является общим местом. Но это не эквивалент энтропийной платы. Утрата упорядоченности — это нечто более сложное, чем трата возобновляемых энергетических ресурсов, и сложнее восстанавливаемое, потому что помимо энергетической включает и информационную составляющую. О том, что это так, наверное, может говорить даже такой простой и общеизвестный факт, что путем повышения одной только энергетики организма далеко не всегда удается справиться с неврологическими патологиями.
Действительно, решение логических задач представляет значительные трудности, которые проявляются в снижении адекватности поведения животных в условии эксперимента. По мнению Л.В. Крушинского, в основе этого явления лежат патологические процессы, развивающиеся из-за перенапряжения мозга, возникающего в результате решения логической задачи. Так, у крыс и черепах в коре и подкорковых структурах мозга после нескольких правильных решений экстраполяционной задачи при использовании метода ЭЭГ была зарегистрирована резко выраженная патологическая активность в виде высоковольтных разрядов и комплексов «пик-волна», характерных для эпилептического припадка. Такие изменения электрической активности предположительно могут быть связаны с нарушением упорядоченных процессов в мозге в результате повышения энтропии в момент решения задачи. Интересно, что, по наблюдениям Л.В. Крушин- ского, эпилептические припадки и срывы в поведении появлялись у «более умных» животных, решающих задачи с первых предъявлений. В то время как «средние» особи, постепенно обучающиеся правильному решению в ходе экстраполяционных проб и ошибок, не проявляли патологий поведения (Семиохина и др., 1976; Крушинский Л. В., 1986).
Вероятно, запас упорядоченности (негэнтропии) должен значительно превышать повышение энтропии, происходящее при решении определенного класса логических задач, с которыми сталкивается животное данного вида. Высокая упорядоченность может быть тем «запасом прочности», который сводит к минимуму негативные последствия (невротические срывы, неадекватное поведение), которые сопровождают процесс мышления. В обеспечении сохранности этого запаса большую роль играют, по-видимому, и биологические (физиологические) механизмы, развившиеся в эволюции для предотвращения срывов и ограничивающие когнитивную активность нервной системы.
С другой стороны, можно предположить, что существование этого «запаса прочности» (определенного уровня упорядоченности, необходимого мозгу для оплаты решения задачи и восстановления) содержит потенциальную возможность потратить этот запас на решение существенно более сложной задачи, очевидно, с утратой стабильности и невозможности полного восстановления упорядоченности нервной системы. О том, что имеющийся запас прочности мозга в ряде случаев может быть необратимо потрачен на решение «более сложных» задач, косвенно свидетельствуют такие явления, как развитие нервно-психических заболеваний у одаренных людей (Ломброзо, 1892).
С точки зрения теории эволюции и естественного отбора можно предположить, что те особи в популяции, которые жертвуют упорядоченностью нервной системы ради решения более сложных задач, оказываются в некотором двояком положении. С одной стороны, они решают задачи, недоступные для остальных, и в этом получают несомненное преимущество, но с другой стороны, теряют оптимальное физиологическое функционирование нервной системы, становясь невротиками и лишаясь ряда преимуществ здоровых особей. Эта двоякость может обеспечить довольно длительное сохранение в поколениях неврологически неустойчивых, но решающих сложные задачи особей. Далее очевидно, что формирование на каком-то этапе эволюции дополнительных ресурсов упорядоченности, необходимых для восстановления нервной системы после решения «задач повышенной сложности», обеспечит этим особям уже несомненное превосходство в борьбе за существование над другими. Очевидно также, что меняющиеся внешние условия будут влиять на соотношение плюсов и минусов наличия склонности к решению более сложных задач. В каких-то условиях наличие этой способности может определить выживание только тех особей, у которых эта склонность была, и тем самым существенно увеличить представленность этого признака для последующего отбора и формирования необходимого компенсаторного усложнения нервной системы.
Таким образом, можно предположить, что само существование определенного запаса упорядоченности, необходимого для компенсации энтропии, возникающей при решении задач, создает условия для вектора эволюции, направленного на усложнение мозга и решение более и более сложных задач. Это предположение не исключает более традиционного подхода, когда повышение сложности и упорядоченности мозга возникает случайно, а затем сохраняется и поддерживается в эволюции как положительный признак, позволяющий животному решить более сложную задачу (Крушинский Л. В., 1974,1977,1986; Шмальгаузен, 1968). Однако более вероятным представляется сценарий, при котором повышенный когнитивный успех сначала оплачивается утратой устойчивого функционирования нервной системы, а уже затем стимулирует поиск компенсаторных решений в эволюции.
Одним из первых биологов, указавших на увеличение упорядоченности нервной системы в процессе эволюции, был выдающийся ученый И.И. Шмальгаузен (1968). По-видимому, эволюционное усовершенствование мозга, позволяющее решать более сложные логические задачи, реализовалось следующим образом. Происходило увеличение общей упорядоченности мозга в результате увеличения количества и разнообразия нейронов и образуемых ими связей, сложности строения самих нейронов, дифференцировки мозга и т. п. Вероятно, также снижалась реальная плата за решение задачи, определяемая неравенством (13), то есть отношение AS / кЛІ уменьшалось в процессе эволюции.
Ряд современных работ приводит рассуждения и свидетельства в пользу растущей упорядоченности мозга в ходе эволюции (Ва$аг et al., 2007; Bayar, Güntekin 2009; Huang, 2008; Friston, 2010; Carhart-Harris et al., 2014). Прежде всего нужно упомянуть теоретические работы, вызвавшие большой резонанс в нейронауке и претендующие на долгожданную общую теорию мозга (Huang, 2008; Fristen, 2010; Fristen et al., 2012 a,b). Эти работы постулируют принцип, в соответствии с которым мозг, как и любая самоорганизующуяся адаптивная система, активно противостоит энтропии (неопределенности), стремясь снизить количество маловероятных состояний и повысить предсказуемость состояний (принцип снижения свободной энергии). Авторы показывают, что практически все виды мозговой деятельности соответствуют этому доминирующему принципу. С ним же, очевидно, согласуются и идеи Дж. Хоукинса о том, что основной функцией мозга является предсказание и формирование внутренних моделей наиболее вероятных внешних конфигураций (Hawkins, Blakeslee, 2005).
Таким образом, разработанная Л.В. Крушинским экстраполяционная теория работы мозга, предполагающая, что животные способны улавливать элементарные законы, связывающие между собой явления природы, приобретает новую актуальность. Характерно в этом контексте название статьи «Животные предвидят наступление событий», посвященной открытию Л.В. Крушинского и вышедшей в 1958 году. Решение когнитивных задач, очевидно, укладывается в обсуждаемый общий принцип, поскольку повышает предсказуемость некоторых внешних событий (причем целого ряда событий в случае, когда речь идет о понимании определенного закона/закономерности).
Трудно не согласиться с тем, что то, каким образом реализуется этот принцип, является наиболее интригующим в понимании работы мозга. В предложенной статье рассматривается вопрос платы за получение новой информации. Я привел теоретические предпосылки и сослался на некоторые экспериментальные данные, свидетельствующие о том, что за получение этой информации мозг должен заплатить временной утратой исходной упорядоченности, которая, по-видимому, не всегда может быть успешно восстановлена за счет повышения энтропии внешней среды. Эти доводы согласуются с флуктуационной теоремой, в соответствии с которой вероятность повышения энтропии в системе со временем выше, чем ее снижение (следствие второго закона термодинамики). О том, что этот момент был упущен в современных представлениях, свидетельствует высказывание того же Фристона: «In other words, biological systems somehow manage to violate the fluctuation theorem, which generalizes the second law of thermodynamics» (Fristen, 2010, p. 127). Наиболее спорным предположением из предложенных здесь для обсуждения является предположение о том, что в эволюции сначала создается некоторый «запас», избыток упорядоченности, которым затем можно оплатить приобретение качественно новой информации.
Литература
Блюменфельд Л.Л. Проблемы биологической физики. М.: Наука, 1974.
Бриллюэн Л. Наука и теория информации. М.: Изд. физмат. лит., 1960.
Бриллюэн Л. Научная неопределенность и информация. М.: Мир, 1966.
Васильев А. М. Введение в статистическую физику. М.: Высшая школа, 1980.
Волькенштейн М.В. Молекулярная биофизика. М.: Наука, 1975.
Волькенштейн М. В. Физика и биология. М.: Наука, 1980.
Волькенштейн М. В. Биофизика. СПб.: Лань, 2008.
Грин Б. Ткань космоса. М.: УРСС, 2000.
Животные предвидят наступление событий. Открытие советского физиолога // Zycie Warszawy. 13.02.1958. №38 (4459).
Зорина 3. А. Мышление животных: эксперименты в лаборатории и наблюдения в природе // Зоологический журнал. 2005. Т. 84. №1.С. 134-148.
Зотин А. И., Зотин А. А. Направление скорость и механизмы прогрессивной эволюции. М.: Наука, 1999.
Кастлер Г. Возникновение биологической организации. М.: Мир, 1967.
Крушинский А. Л. Биофизические аспекты рассудочной деятельности // Формирование поведения животных в норме и патологии. К 100-летию со дня рождения Л. В. Крушинского (1911-1984) / Под ред. И. И. Полетаевой, 3. А. Зориной. М.: Языки славянской культуры, 2013. С. 424-436.
Крушинский Л. В. Элементарная рассудочная деятельность животных и ее роль в эволюции // Философия в современном мире. Философия и теория эволюции. М.: Наука, 1974. С. 156-215.
Крушинский Л. В. Биологические основы рассудочной деятельности. М.: Изд. МГУ 1977.
Крушинский Л. В. Биологические основы рассудочной деятельности. 2-е изд., перераб. и доп. М.: Изд. МГУ, 1986.
Крушинский Л. В. О возможном механизме рассудка // Формирование поведения животных в норме и патологии. К 100-летию со дня рождения Л. В. Крушинского (1911 -1984) / Под ред. И. И. Полетаевой, 3. Зориной.М.: Языки славянской культуры, 2013. С. 437-450.
Ленинджер А. Основы биохимии. Т. 2. М.: Мир, 1985.
Ломброзо Ч. Гениальность и помешательство. СПб.: Издание Ф. Павленкова, 1892.
Перепелкина О. В., Маркина Н. В., Полетаева И. И. Способность к экстраполяции направления движения у мышей, селектированных на большой и малый вес мозга: влияние пребывания в «обогащенной» среде // Журнал высшей нервной деятельности им. И. П. Павлова. 2006. Т. 56. №2. С. 244-248.
Полетаева И. И. Гипотеза Л. В. Крушинского «О возможном механизме рассудка» и развитие нейробиологии во второй половине XX века // Формирование поведения животных в норме и патологии. К 100-летию со дня рождения Л. В. Крушинского (1911-1984) / Под ред. И. И. Полетаевой, З.А. Зориной. М.: Языки славянской культуры, 2013. С. 451-454.
Попова Н. В., Кесарев В. С., Полетаева И. И., Романова Л. Г. Корковая цитоархитектоника корковых структур у мышей, селектированных на большой и малый вес мозга // Журнал высшей нервной деятельности им. И. П. Павлова. 1983. Т. 33. №3. С. 576-582.
Пригожин И., Стенгере И. Порядок из хаоса. М.: УРСС, 2000.
Романовский Ю. М., Степанова Н. В., Чернавский Д. С. Математическая биофизика. М.: Наука, 1984.
Рубин А. Б. Биофизика. Книга 1. Теоретическая биофизика. М.: Высшая школа, 1987.
Сахаров Д. А. Биологический субстрат генерации поведенческих актов // Журнал общей биологии. 2012. Т. 73. №5. С. 334-348.
Семиохина А. Ф., Очинская Е. И., Рубцова Н. Б., Крушинский Л. В. Новое в изучении экспериментальных неврозов, вызванных перенапряжением высшей нервной деятельности // Доклады АН СССР. 1976. Т. 231. №2. С. 503-505.
Тринчер К. С. Биология и информация. М.: Наука, 1965.
Хакен Г. Информация и самоорганизация. М.: Мир, 1991.
Чернавский Д. С. Синергетика и информация. М.: УРСС, 2004.
Шмальгаузен И. И. Кибернетические вопросы биологии. Новосибирск: Наука, 1968.
Шредингер Э. Что такое жизнь с точки зрения физики. М.: Гос. изд-во иностранной литературы, 1947.
Эбелинг В., Энгель А., Файстель Р. Физика процессов эволюции. М.: УРСС, 2001.
Bargmann С. I. Beyond the connectome: how neuromodulators shape neural circuits // Bioessays. 2012. Vol. 34. No. 6. P. 458-465. doi: 10.1002/bies.201100185
Bafar E., Güntekin В. A breakthrough in neuroscience needs a “Nebulous Cartesian System”: Oscillations, quantum dynamics and chaos in the brain and vegetative system // International Journal of Psychophysiology. 2007. Vol. 64. No. 1. P. 108-122. doi:10.1016/j.ijpsycho.2006.07.012
Bafar E., Güntekin В. Darwin's evolution theory, brain oscillations, and complex brain function in a new “Cartesian view” // International Journal of Psychophysiology. 2009. Vol. 71. No. 1. P. 2-8. doi:10.1016/j.iipsycho.2008.07.018
Brillouin L. Physical entropy and information. II // Journal of Applied Physics. 1951. Vol. 22. No.3. P. 338-343. doi:10.1063/l.1699952
Carhart-Harris R. L., Leech R., Hellyer P. J., Shanahan M., Feilding A., Tagliazucchi E., Chialvo D. R., Nutt D. The entropic brain: a theory of conscious states informed by neuroimaging research with psychedelic drugs // Frontiers in Human Neuroscience. 2014. Vol. 8. doi:10.3389/fhhum.2014,00020
Davie C.A. A review of Parkinson's disease 11 British Medical Bulletin. 2008. Vol. 86. No.l. P. 109-127. doi:10.1093/bmb/ ldn013
Fristen К. The free-energy principle: a unified brain theory? // Nature Reviews Neuroscience. 2010. Vol. 11. No. 2. P. 127-138. doi:10.1038/nrn2787
Fristen К., Breakspear M., Deco G. Perception and self-organized instability // Frontiers in Computational Neuroscience. 2012a. Vol. 6. P. 44. doi:10.3389/fncom.2012.00044
Fristen К., Thornton C., Clark A. Free-energy minimization and the dark-room problem // Frontiers in psychology. 2012b. Vol. 3. P. 130. doi:10.3389/fpsyg.2012.00130
Hawkins J., Blakeslee S. On intelligence. New York: Times Books, 2004.
Huang G. Is this a unified theory of the brain 11 New Scientist. 2008. Vol. 2658. P. 30-33.
Toga A. W., Clark K. A., Thompson P. M., Shattuck D. W., Van Horn J. D. Mapping the human connectome 11 Neurosurgery. 2012. Vol. 71. No. 1. P. 1-5. doi:10.1227/NEU.0b013e318258e9ff
Zecevic D., Wu J.-Y, Cohen L. B., London J., Hopp H, Falk С. X. Hundreds of neurons in the Aplysia abdominal ganglion are active during the gill-withdrawal reflex 11 The Journal of Neuroscience. 1989. Vol. 9. No. 10. P. 3681-3689.
Related Articles
Андрей Курганский
Яницкий Олег Николаевич
Красовская Людмила Владимировна
Исабекова Тамила Илахидиновна
Михайлов Андрей Валерьевич
