This article is published under a Creative Commons license and not by the author of the article. So if you find any inaccuracies, you can correct them by updating the article.

Remote sensing, snow modelling, survey data and Indigenous Knowledge show how snow and sea-ice conditions affect Peary caribou (Rangifer tarandus pearyi) distribution and inter-island and island–mainland movements



Coralie Gautier
Alexandre Langlois
Vincent Sasseville
Erin Neave
Cheryl Ann Johnson
Published: Jan. 1, 2022
Latest article update: July 26, 2023
This article is published under the license




Abstract
Accelerated warming of the Arctic has reduced sea ice and has increased the occurrence of winter extreme events like rain-on-snow and storms that impact snow-cover densification, affecting Peary caribou (Rangifer tarandus pearyi) seasonal movements and grazing conditions. We used caribou movements between Banks, Melville and Victoria islands and mainland Canada, documented from Indigenous Knowledge, to assess whether spatiotemporal trends in sea-ice anomalies (1983–2019) can be used as an indicator of caribou movement. We used the SNOWPACK model to evaluate how foraging conditions (as indexed by simulated snow properties) contribute to the prediction of caribou presence. Our results suggest that changes in sea-ice anomalies over time have impacted caribou crossings between islands: caribou no longer use areas with less sea ice whilst they continue to use areas with more sea ice. Our model evaluation shows that, when the simulated snow conditions are paired with other environmental variables, the ability of models to predict Peary caribou occurrence on land was enhanced across Banks and Melville islands. Overall, the land models suggest that caribou are more likely to occupy areas with lower density of snow accumulation and a majority of forb tundra with dwarf shrubs for Banks Island and cryptogam tundra, rush and grass for the Melville Island Complex. Our results suggest that future work monitoring changes in sea-ice and snow conditions will be important for understanding the impact of climate change on the distribution of Peary caribou in the western Arctic.
Keywords
Climate change, Arctic amplification, seasonal movement, snow densification
Introduction
Observed changes in climate and climate variability are significantly impacting the Arctic environment, where warming is occurring twice as fast as elsewhere on the planet (Rinke & Dethloff 2008; Liston & Hiemstra 2011; Uboni et al. 2016). This acceleration is known as Arctic amplification, a combination of positive feedback loops that create faster warming (Serreze & Barry 2011; Uboni et al. 2016; Langlois et al. 2017), with consequences that include an increase in the frequency and intensity of extreme weather events (Il Jeong & Sushama 2018; Vincent et al. 2018). More specifically, forecasting models and recent remote sensing techniques show an increase in the frequency of ROS events (Liston & Hiemstra 2011; Dolant et al. 2017; Sobota et al. 2020), and a general increase in total precipitation (Hansen et al. 2014; Johnson et al. 2016). The general trend towards warming has caused a reduction in winter snow-cover duration, especially during spring (Callaghan et al. 2011; Derksen & Brown 2012; Bokhorst et al. 2016), a retreat of glaciers (Gardner et al. 2012; Papasodoro et al. 2015) and a decrease in sea-ice cover with a higher incidence of polynyas (Serreze & Stroeve 2015; Stroeve & Notz 2018; Cai et al. 2021).
These phenomena have had a significant impact on snow conditions, more specifically on the snowpack stratigraphy (Bokhorst et al. 2016; Dolant et al. 2017; Langlois et al. 2017). Warming temperatures, sustained winds and ROS events have led to densification of snow cover in the polar regions (Liston & Hiemstra 2011; Dolant et al. 2016; Royer et al. 2021). More specifically, Ouellet et al. (2017) demonstrated that the densification of the snow and its temporal sustainability (cumulated thickness of snow above a density threshold of 300 to 400 kg∙m–3) has an impact on caribou (Rangifer tarandus) population numbers. This was later confirmed by Langlois et al. (2017) and Dolant et al. (2018), who highlighted the impact of storms and ROS events on caribou population numbers and mortality, respectively. For example, mass caribou mortality events have been associated with severe weather, ROS and sea-ice melt in different regions of the Arctic (Parker et al. 1975; Kohler & Aanes 2004; Tews et al. 2007; Forbes et al. 2016; Dolant et al. 2018). This is problematic from a population dynamics perspective, as well as cultural perspective since caribou are an important source of food to local communities (Ljubicic et al. 2018; Tomaselli et al. 2018).
Peary caribou (Rangifer tarandus pearyi) are endemic to the Canadian Arctic Archipelago and live in small groups of 2–15 animals (COSEWIC 2015). This northernmost caribou Designable Unit is listed as Threatened by the Committee on the Status of Endangered Wildlife in Canada (COSEWIC 2015) on the account of widespread population declines observed over the last three caribou generations (i.e., 35% decrease in 27 years; COSEWIC 2015).
Sea ice is a key habitat for Peary caribou. It is of particular importance for seasonal movements to areas with more accessible or higher quality forage and lower predation risk (Johnson et al. 2016). Current warming trends have led to a reduction of both first-year and multi-year sea ice, along with significant inter-annual variation in distribution and concentration (Serreze & Francis 2006; Stroeve et al. 2007; Howell et al. 2009; Holland et al. 2010; Serreze & Stroeve 2015; Blanchard-Wrigglesworth et al. 2011; Bushuk & Giannakis 2017). During the period of 1968–2016, summer sea-ice cover in the Canadian Arctic Archipelago decreased by 4.8% per decade (Mudryk et al. 2018; Derksen et al. 2019). Changes in the spatial and temporal distribution of sea ice impact Peary caribou movement between islands of the Canadian Arctic Archipelago (Stien et al. 2010; Jenkins et al. 2016; Le Corre et al. 2017), particularly when sea ice is thin or moving fast. Indeed, the deterioration of sea-ice conditions during seasonal movement has resulted in caribou mortality (Parker et al. 1975; Tyler 2010). Because sea-ice conditions are forecasted to change even more in the future (Guarino et al. 2020), an assessment of how current and future changes might influence Peary caribou is needed to guide recovery efforts (ECCC 2021).
Recent advances in snow models such as SNOWPACK (Bartelt & Lehning 2002; Lehning et al. 2002) have shown promise in addressing research gaps regarding the effects of climate change or, more specifically, changing snow conditions on caribou/reindeer (Vikhamar-Schuler et al. 2013; Turunen et al. 2016; Ouellet et al. 2017; Boelman et al. 2019; Martineau et al. 2022). Species distribution models are commonly used to estimate the relative importance of different environmental variables with respect to the distribution of wildlife when occurrence data are sparse or limited in coverage (Elith & Leathwick 2009 ; Elith et al. 2011; Smith & Santos 2020). Johnson et al. (2016) used a species distribution model called MaxEnt to model factors influencing the spatial distribution of Peary caribou by linking location data from surveys and telemetry and participatory mapping and interviews with Inuit and Inuvialuit to relevant environmental data (including vegetation, elevation, snow, precipitation, wind and sea-ice fraction). The resolution (25 km) and uncertainty in the snow data were identified as a limitation in the analysis by the authors. Similarly, other studies examined the importance of snow to Peary caribou in different areas of the Canadian Arctic Archipelago using snow data with resolutions of >10 km (Ouellet et al. 2017; Jenkins et al. 2020; Kaluskar et al. 2020; Martineau et al. 2022). None of these studies examined how changes in sea-ice conditions might influence seasonal movement between islands, despite the significance of sea ice for Peary caribou survival. Jenkins et al. (2016) highlighted how deteriorating sea-ice conditions could cause caribou to swim across the sea or take detours or, ultimately, create a barrier to movement altogether.
Our paper aims to improve knowledge of the influence of sea-ice reduction and snow densification on seasonal movements and distribution of Peary caribou. More specifically, we: (1) evaluate whether spatiotemporal changes in young sea-ice concentration between 1983 and 2019 along three inter-island crossings around Banks Island adequately capture changes in caribou movement patterns described by Inuit and Inuvialuit Knowledge; (2) use OSSA (Ouellet et al. 2017) to simulate snow conditions affecting foraging (i.e., daily cumulative thickness of snow over 350 kg/m³ of density [CT350]; Vikhamar-Schuler et al. 2013; Ouellet et al. 2017) at a finer resolution (4 km) than previous work; and (3) model seasonal distributions of Peary caribou on land using the finer resolution snow conditions identified in (2), elevation and land cover types, with the MaxEnt predictive software.
Data and methods
Study site
The study area was located in the Northwest Territories (Canada), around Banks Island and the Melville Island Complex, which is the name given to the group of islands including Melville, Byam-Martin, Mackenzie-King, Brock and Borden islands (Fig. 1). Snow conditions were modelled across all islands for the analyses of Peary caribou occurrence.
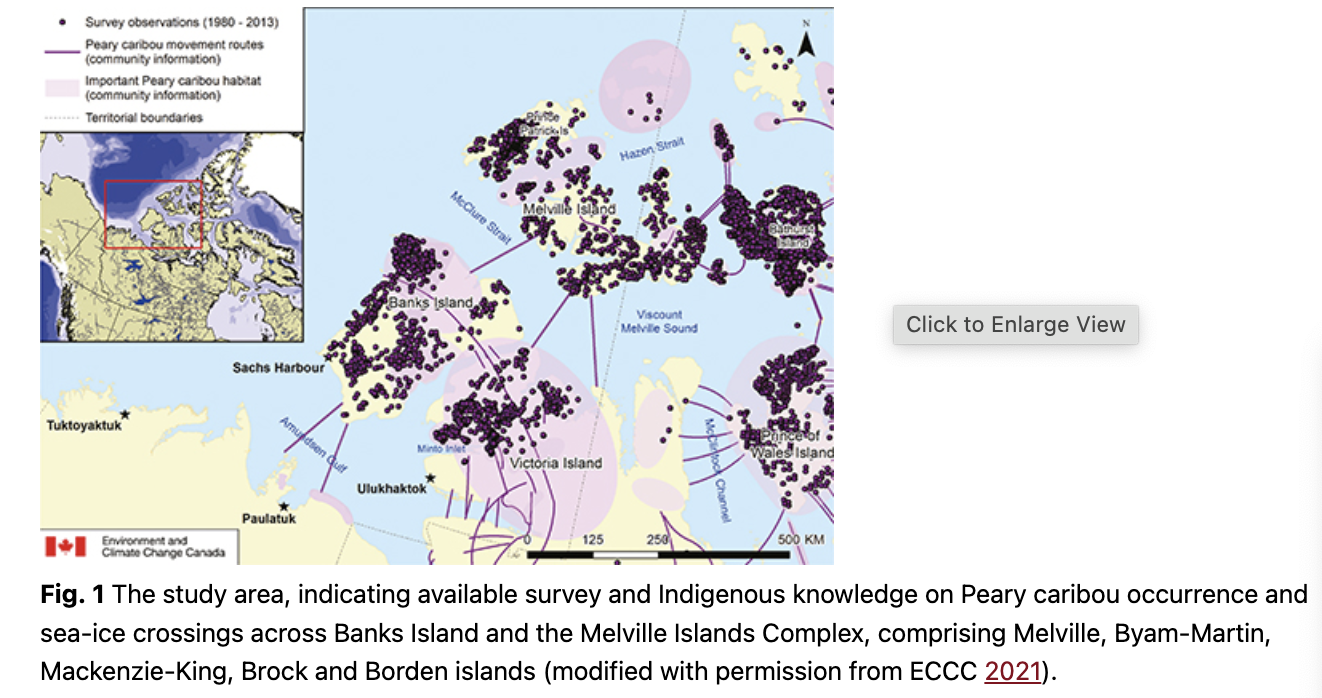
Three zones surrounding Banks Island were identified for the analysis of the trends in sea-ice anomalies (Fig. 2). These zones were selected based on Inuvialuit and Inuit Knowledge of Peary caribou sea-ice crossings. Johnson et al. (2016) provide a description of the community workshops and methods for weaving Inuit/Inuvialuit Knowledge with Western science used in this study.
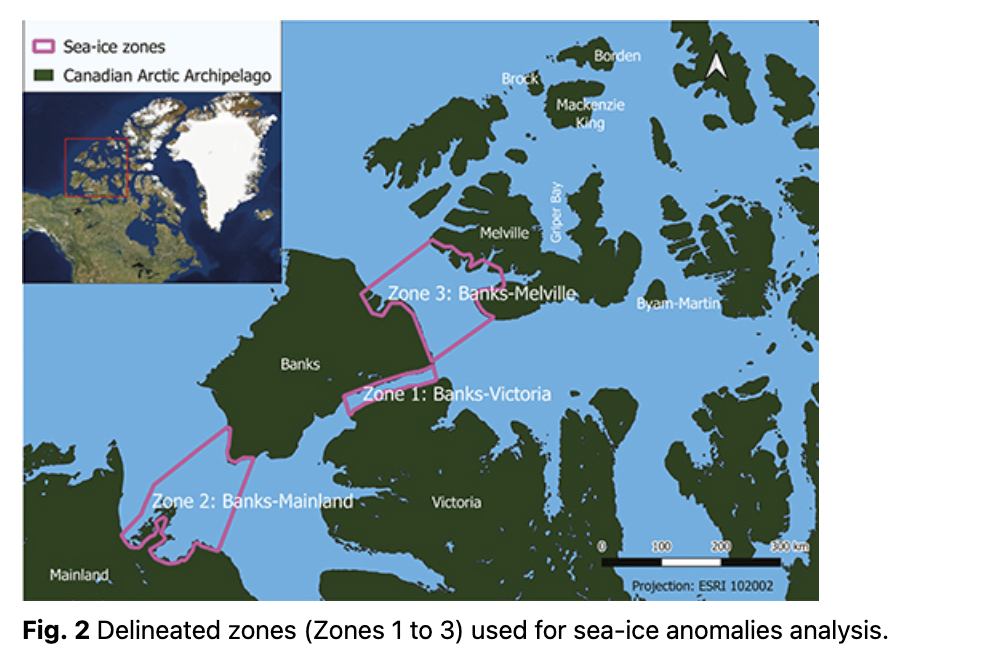
Zone 1 is located between Banks Island and Victoria Island, a 15–20 km crossing zone (area of 3600 km²). Caribou used this crossing to move between the islands throughout the study period, i.e., 1983–2019 (Olohaktomiut HTC 2013; Sachs Harbour HTC 2013).
Zone 2 is located between Banks Island and the mainland, an approximate distance of 130–150 km (area of 24 600 km²). Caribou used this zone at the beginning of the study period (1980s and 1990s) but have since stopped using it (Paulatuk HTC 2013).
Zone 3 is located between Banks Island and Melville Island, a distance of 100–110 km (area of 23 000 km²). Caribou tracks are observed when people travel to this area about every 4 yr, but there is a desire to know whether conditions might be suitable for crossing because there have been fewer observations of caribou crossing in the area (Paulatuk HTC 2013; Olohaktomiut HTC 2016; Sachs Harbour HTC 2016).
Peary caribou are generally described as moving south in the fall, including movement along the east side of Victoria Island and from Victoria Island onto the mainland, and moving back north in the spring (Olohaktomiut HTC 2013; Paulatuk HTC 2013; Sachs Harbour HTC 2013).
Sea ice
We used the different ice types defined by the Canadian Ice Service Digital Archive (Government of Canada 2020c) to calculate seasonal sea-ice concentration anomalies, following the methodology proposed by Couture (2022). The digital archive provides weekly sea-ice charts for the western Arctic from 1968 onwards based on available satellite imagery, weather and oceanographic information, and visual observations (Tivy et al. 2011). The sea-ice concentration polygons in the database are of variable size, ranging 2–5 km² to >100 km². Satellite data contributed to 50–55% of all available data between 1978 and 1996 and up to >80% after 1996 with the launch of RADARSAT-1 (Tivy et al. 2011; Couture 2022). We used the digital archive data from 1983 onwards given changes in technology and methods over time, interpolation issues arising from few available images (Tivy et al. 2011).
Couture (2022) developed a methodology to assess the sea-ice conditions in the Canadian Arctic Archipelago, focusing on the development of a polynya climatology using sea-ice concentration (% of cover). To quantify sea-ice concentration and detect open water and opening periods, a threshold for sea-ice conditions was used to extract open water and young ice areas from the Canadian Ice Service Digital Archive that are representative of polynyas. Polygons within the ice chart that had an ice concentration <70% were designated ‘open water,’ in accordance with the Manual of ice (Canadian Ice Service 2005). This was also verified by comparing polygons with less than 70% ice concentration with MODIS TERRA and AQUA imagery for the corresponding ice chart dates. During this phase, areas with seemingly thinner ice polygons with ice concentration above 70% were also selected if the ice within them consisted mainly of the younger ice types such as new ice, grey ice, grey-white ice and first-year thin ice (Joint Technical Commission for Oceanography and Marine Meteorology 2014), so that an area of open water surrounding the islands was quantified.
On the basis of Indigenous Knowledge, Paquette (2020) determined that 10 cm is the minimum thickness that caribou can walk on. Based on that, we applied Couture’s methods (2022), focusing on ‘young ice’ only, and using the data from the Canadian Ice Service Digital Archives (Government of Canada 2020c), we combined the following categories of ice concentration: ‘first-year ice,’ ‘first-year thick ice,’ ‘first-year medium ice,’ ‘first-year thin ice,’ ‘young ice,’ ‘grey-white ice’ and ‘grey ice’ into a new layer called ‘total young ice’ concentration. The new layer was converted from a vector to a raster dataset summarizing the percentage of ‘total young ice’ in each pixel (500 × 500 m). This new raster layer was then used to calculate yearly anomalies (difference between the concentration value of 1 year and the mean concentration value of all years) in total young ice concentration between 1983 and 2019 for each Peary caribou season. Positive anomalies mean more ice in a given year compared to the average of all years and vice versa for negative anomalies.
We divided the sea-ice data into pre- and post-2000 on the basis of reductions in the extent, thickness and volume of sea ice reported in the literature in the 2000s compared to the 1970s and 1980s (Stocker et al. 2013). We compared the long-term averages in anomalies across the three zones of interest over the period (1983–2019), as well as differences in the magnitude of anomalies between the pre- and post-2000 periods using repeated measures ANOVA adjusted for unequal sample sizes. We only report significant (p ≤ 0.05) or marginally significant results (p < 0.100).
Snow state variables
The SNOWPACK model simulates each layer of the snowpack with its bulk (thickness, water equivalent and mass balance) and layered physical characteristics (microstructure, density, temperature, etc.; Bartelt & Lehning 2002; Lehning et al. 2002). Ouellet et al. (2017) created OSSA to facilitate wider use of the SNOWPACK model at different spatial resolutions (Beaudoin-Galaise 2016). This version includes the Groot Zwaaftink et al. (2013) Antarctic version of SNOWPACK that improves snow-density simulations for the Arctic by changing the way the model handles snow deposition (density up to 600 kg/m³). It also allows snow to accumulate only above a wind-speed threshold of 4 m/s (Martineau et al. 2022). OSSA includes a density correction for the basal layers proposed by Gouttevin et al. (2018), where density is fixed at 150 kg/m³ until snow height reaches vegetation height, to model more precisely the Arctic snow conditions. Martineau et al. (2022) highlighted the necessity of integrating the last physical corrections derived by Gouttevin et al. (2018) to SNOWPACK to better represent the snow characteristics in the Arctic.
A variety of different data sets were used as input for OSSA, including land cover (for the vegetation albedo), topography (for the slope angle and the altitude) and meteorological input variables (Ouellet et al. 2017). Meteorological variables include air and ground temperature, relative humidity, wind speed, short wavelength incident rays, long wavelength incident rays and precipitation (Vikhamar-Schuler et al. 2013; Turunen et al. 2016; Ouellet et al. 2017) from NARR and from the National Centers for Environmental Prediction Environmental Modeling Center (NOAA 2020). The horizontal resolution of NARR is 0.3 degrees (approximately 32 km), with a temporal resolution of eight times daily (every 3 hours).
The spatialized snow simulations can be run on different spatial resolutions, where finer scales are simulated solely on the basis of soil properties in ‘column’ mode (i.e., simulated vertically for each grid, with no adjacency effects). Soil properties include topography and soil albedo at 100 m spatial resolution and averaged over 4 × 4 km to match the snow simulations; Beaudoin-Galaise (2016) showed that these properties improve local representation of bulk snow properties like depth and snow-water equivalent. The land cover information was extracted from circa 2000, a vector data set (Government of Canada 2020b) for which the soil roughness values (Meloche et al. 2020) and the thermal conductivity values (Sasseville 2020, unpubl. data) were associated with each class of the land cover. Finally, topographic information was retrieved from the Canadian Digital Elevation Model (Government of Canada 2020a) raster with a spatial resolution of 100 m (i.e., elevation, slope orientation and slope angle).
OSSA allows for spatialization of the snow simulation outputs. In this study, we chose to use the cumulative thickness in cm of snow layers surpassing the defined critical density value of 350 kg/m³ (CT350) as a predictor of caribou occurrence. This density appears to hinder reindeer and caribou foraging, resulting in starvation (see Vikhamar-Schuler et al. 2013; Ouellet et al. 2017). We simulated the CT350 for the years 2000 to 2013 (resolution of 4 km), over Banks Island and the Melville Island Complex and calculated an average for the entire period, to match the caribou location data.
MaxEnt fitting and caribou data from surveys and Indigenous knowledge
MaxEnt is a software used to assess species’ distributions (Phillips & Dudík 2008; Ahmed et al. 2015; Phillips et al. 2019). The model uses ‘maximum entropy’ or the probability of the distribution having the widest, almost uniform distribution (Phillips et al. 2006). MaxEnt aims to identify the most influential environmental variables that explain probability of occurrence or presence (Elith et al. 2011). The use of MaxEnt has proved useful for defining the occurrence of a variety of plant and animal species (Elith et al. 2006; Elith et al. 2011) and is well suited to our Peary caribou data, given that the available data are ‘presence-only’: no data could mean either that there was no caribou or that the area was never surveyed (Elith et al. 2011).
Three major biological seasons were identified to reflect changes in factors affecting Peary caribou habitat selection: (1) spring movement and calving (April–June, hereafter ‘spring’); (2) summer foraging and rut (July–October, hereafter ‘summer–fall’); and (3) winter movement and foraging (November–March, hereafter ‘winter’). During calving, parturient females choose habitats to reduce calf predation risk over high-quality foraging areas (Johnson et al. 2016). During summer–fall, caribou search for habitats with abundant food resources to store fat reserves for the leaner winter months (Johnson et al. 2016). During winter, unfavourable climatic conditions can limit access to food (Johnson et al. 2016), which can result in starvation and death (e.g., Dolant et al. 2018).
We used three explanatory variables in our MaxEnt models (version 3.4.1): cumulative snow thickness (cm) over 350 kg/m³ from OSSA; vegetation types based on the Circumpolar Arctic Vegetation Map (CAVM Team 2003); and elevation. We used data from Johnson et al. (2016) that included geographically referenced locations of Peary caribou collected from governmental aerial and terrestrial surveys (conducted from 2000 to 2013), and community observations mapped during workshops in the communities of Taloyoak, Ulukhaktok, Sachs Harbour (Ikahuak), Cambridge Bay (Iqaluktuttiaq), Kugaaruk, Resolute Bay (Qausuittuq), Grise Fiord (Aujuittuq), Paulatuk and Gjoa Haven (Uqsuqtuuq). Caribou locations were assigned to the appropriate biological season agreed upon during the workshops.
All environmental data were formatted to the same extent, projection and grid size (4 × 4 km) for MaxEnt. We ran seven models (all possible combinations using vegetation, snow and elevation as explanatory variables) for each season across Banks Island and the Melville Island Complex. We could model the winter distribution of caribou only on the Melville Island Complex because of the lack of caribou observations on Banks Island during this season. We used AICc to evaluate model performance and identify the model that best explained the distribution of caribou across each of the islands (Portet 2020). We used the AUC to further evaluate how well the model predicted Peary caribou locations. Typically, an average AUC value >0.700 is considered to predict better than random according to Elith et al. (2006). We used an eightfold cross-validation procedure on the model with the three variables (CT350, land cover, elevation) to assess if it was better than the random test point method where 20% of the observations were withheld for model validation. Since both methods showed similar statistical results, we report the second method only. We used permutation importance, which represents the amount of information gained or explained by one variable compared to what would be explained if the variable was random, to assess relative variable importance.
Results
Sea ice
Spring. The positive sea-ice anomalies in the spring mean anomalies map for 1983–2019 (mean of all the annual anomalies, pixel by pixel; Fig. 3a) suggested a global increase of young sea-ice concentration over time. There was a tendency for spring sea-ice anomalies to differ between the zones (ANOVA: p = 0.084), with a few negative sea-ice anomalies (decrease of young sea-ice concentration) between Banks Island and the mainland (Zone 2) and Banks Island and Melville Island (Zone 3) but none between Banks Island and Victoria Island (Zone 1).
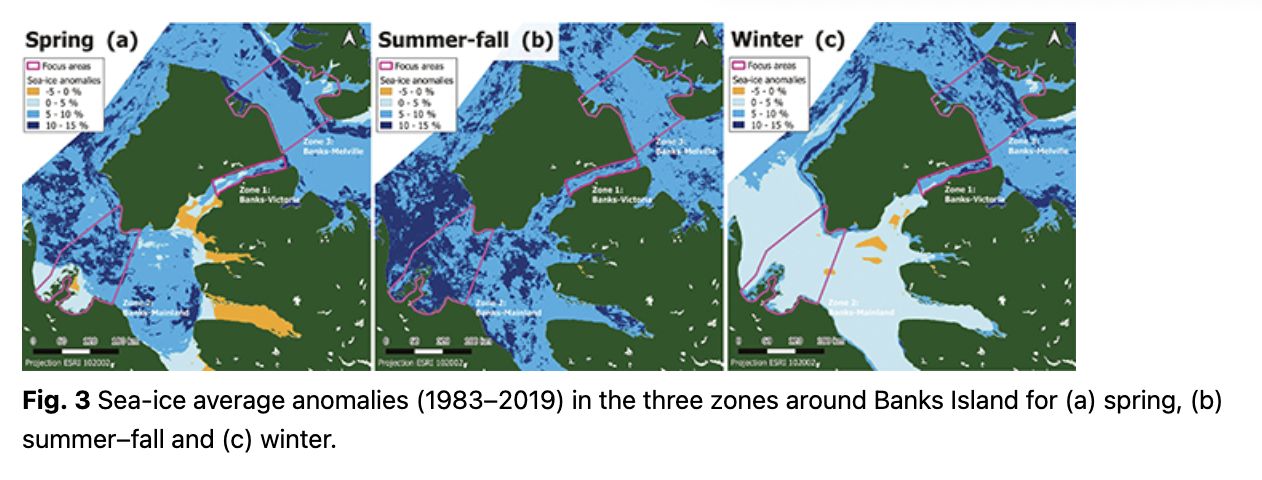
Changes in spring sea-ice anomalies between 1983–1990 and 2000–2019 tended to differ amongst the zones (time by zone interaction: p = 0.070). Zone 2 appeared to experience a decrease in the concentration of first-year ice between 2000 and 2019 compared to the past (Table 1). In contrast, Zones 1 and 3 appeared to experience increases in young sea-ice concentration relative to the long-term average since 2000 (Table 1).
Table 1 Spring, summer–fall and winter sea-ice mean anomaly and concentration before and after 2000 for the three zones around Banks Island. Winter values are not reported because no temporal trends were detected. | ||||
Sea-ice mean anomalies of concentration | Sea-ice mean concentration | |||
Spring | 1983–1999 | 2000–2019 | 1983–1999 | 2000–2019 |
Zone 1 | 2.9% | 9.7% | 1.4% | 2.8% |
Zone 2 | 11.1% | 6.4% | 12.8% | 9.8% |
Zone 3 | 7.2% | 8.6% | 6.9% | 8.0% |
Summer–fall | ||||
Zone 1 | 14.0% | 3.0% | 7.7% | 4.8% |
Zone 2 | 8.0% | 9.3 | 14.9% | 15.5% |
Zone 3 | 13.4% | 7.6% | 13.4% | 8.9% |
Summer–fall. Similar to spring, average sea-ice conditions differed between the zones, but Zone 2 had the highest positive anomalies or increases in young sea-ice concentrations compared to the other zones for summer–fall (p = 0.003; Fig. 3b). There was an overall decrease in the concentration of young sea ice relative to the long-term average when comparing the 1983–1999 period to 2000–2019 (p = 0.050; Table 1) across all three zones. However, the magnitude of the effect differed amongst them (time by zone interaction: p < 0.005) with the decrease being most pronounced between Banks Island and Victoria Island (Zone 1) and Zone 2 showing a slight positive increase in young sea-ice concentrations in summer–fall (Table 1).
Winter. Average sea-ice anomalies differed between the zones (p < 0.005; Fig. 3c). All sea-ice anomalies were positive, indicating a global increase in the concentration of young sea ice. Zone 3, between Banks Island and Melville Island, experienced the most pronounced increase in winter sea-ice anomalies, whereas Zone 2, between Banks Island and the mainland, experienced the least (Fig. 3c). There were no detectable temporal trends in winter sea-ice anomalies from 1983 to 2013 (p ≥ 0.20).
OSSA snow simulations
Spring. OSSA simulations during spring suggested that CT350 values were low (<7000 cm) over more than half of the spatial extent of the islands (Fig. 4a). Areas showing the lowest CT350 (0–7000 cm) were located on the northern and eastern portions of Banks Island, eastern Melville, Byam-Martin, Brock, northern Borden and eastern Mackenzie-King islands. Areas with the highest CT350 (50 000–70 000 cm) values were located on the south coast of Banks Island, and, on Melville Island, on the west and south coasts of Melville Island and the east shoreline of Hecla and Griper Bay. The average for spring was 9215 ± 8291 cm.

Summer–fall. CT350 was extremely low in summer–fall (725 ± 293 cm; Fig. 4b) relative to the other seasons, likely because of periods of no snow. Most values occurred between 400 cm and 800 cm. Areas with the lowest CT350 values (0–400 cm) occurred inland on all the islands. The highest CT350 values (1200–2000 cm) occurred primarily in the northern portion of the Melville Island Complex (i.e., Melville, Mackenzie King and Borden islands).
Winter. Average snow density was highest in winter (13 050 ± 11 265 cm), but both islands had relatively low CT350 values in winter (0–7000 cm; Fig. 4c). These areas covered approximately 50% less of Banks Island compared to spring. Eastern Banks, east-central Melville, Byam-Martin, Brock, northern Borden and eastern Mackenzie–King islands had the lowest CT350 (0–7000 cm) values. Areas with the highest CT350 values (60 000–70 000 cm) were on the south of Banks Island, and Melville Island’s west coast and east shoreline of Hecla and Griper Bay.
Caribou occurrence
Top models differed between the islands and across seasons (Table 2; Supplementary Table S1),but almost all models included snow density or CT350 as a predictor of Peary caribou occurrence, with the exception of the Melville Island Complex in summer–fall.
Table 2 Top seasonal models produced from MaxEnt predicting the probability of Peary caribou occurrence across Banks Island and the Melville Island Complex from 2000 to 2013. AICc values are provided in Supplementary Table S1. | |||||
Model | Banks spring Model 7 | Banks summer–fall Model 7 | Melville spring Model 1 | Melville summer–fall Model 6 | Melville winter Model 7 |
AUC | 0.70 | 0.63 | 0.76 | 0.57 | 0.79 |
AUC SD | 0.04 | 0.04 | 0.06 | 0.02 | 0.03 |
CT350 | 82.7% | 55.6% | 15.5% | – | 47.6% |
Elevation | – | – | 79.6% | – | – |
Vegetation | 17.3% | 44.4% | 4.9% | 100% | 52.4% |
Spring. Snow density (CT350) and vegetation best explained caribou occurrence on Banks Island during spring (model 7; Fig. 5a). The average model AUC (0.704 with SD of ± 0.039; Table 2) exceeded the recommended threshold of 0.7 by Elith et al. (2006) to discriminate presences from random locations. The permutation importance indicated that CT350 (82.7%) provided more explanatory power about caribou occurrence than vegetation (17.3%; Table 2), with Peary caribou preferentially occupying areas of low CT350 (Supplementary Fig. S1a). Sedge/grass, moss wetland was the predominant land-cover class occupied by caribou on Banks Island, followed by the prostrate dwarf-shrub, herb tundra (Supplementary Fig. S1b).

The top model for the Melville Island Complex included elevation as well as CT350 and vegetation (model 1; AUC 0.763 ± 0.058). Elevation played a role in predicting caribou occurrence on this island in spring with a permutation importance of 79.6% compared to 15.5% for CT350 and 4.9% for vegetation (Table 2). Peary caribou preferentially occupied areas of low elevation (0–100 m), low CT350 (<7000 cm on a maximum of 70 000 cm) and cryptogam, herb barren and rush/grass, forb cryptogram tundra land-cover types (Supplementary Fig. S1, e-f-g).
Summer–fall. CT350 and vegetation best explained caribou occurrence on Banks Island during summer–fall (model 7; Fig. 5b), similar to the spring model. However, the AUC value (0.632 ± 0.036) indicates that the model’s ability to discriminate Peary caribou presence from random locations was lower than that of the model for spring. Both variables made similar contributions to the model (55.6% for CT350 and 44.4% for vegetation; Table 2). Areas with high occurrence probability were associated with low CT350 (0–600 cm on a maximum of 2000 cm) and a land-cover type of sedge/grass moss wetland, followed by three other vegetation types: cryptogam, herb barren; graminoid, prostrate dwarf-shrub, forb tundra; low-shrub tundra (Supplementary Fig. S1c–d). In contrast, areas with lower probability of occurrence were associated with non-tussock sedge, dwarf-shrub moss tundra (Supplementary Fig. S1d).
Predicted occurrence values on the Melville Island Complex were higher (60–80%) compared to those on Banks Island, which fell mainly in the 40–60% range (Fig. 5b). For the Melville Island Complex, the top model included vegetation only in this season (model 6). Areas with a higher probability of occurrence were associated with four vegetation types (rush/grass, forb and cryptogam tundra; cryptogam herb barren; cryptogam barren complex (bedrock); prostrate/hemiprostrate dwarf-shrub tundra), whereas areas with low probability of occurrence were associated with the carbonate and noncarbonate mountain complex land cover type (Supplementary Fig. S1h). Overall, the AUC of the model for this season on Melville (0.574 ± 0.021) indicated poor model performance.
Winter. Caribou occurrence on the Melville Island Complex was best explained by CT350 and vegetation in winter (model 7). Both variables appeared to have equal contributions with CT350 at 47.6% and 52.4% for vegetation (Table 2). The AUC value of the model was 0.787 (± 0.025), representing the highest AUC of all models tested. The probability of caribou presence across the Melville Island Complex was quite low in winter, with most values falling below 20% (Fig. 5c). Caribou were more likely to occur in areas of rush/grass, forb, cryptogam tundra or cryptogam, herb barren and CT350 between 7000 and 20 000 cm (with a maximum of 70 000 cm; Supplementary Fig. S1i–j).
Discussion
Sea-ice anomalies
Our results indicate that, on average, positive sea-ice anomalies have predominated around Banks Island, indicating higher than average young sea-ice concentration since 1983. This result may seem odd given studies describing the loss of sea ice in the Arctic (Rigor & Wallace 2004; Maslanik et al. 2011; Yadav et al. 2020; Cai et al. 2021). However, our study encompasses one region of the Arctic and focuses solely on young sea ice. Multi-year ice concentration is decreasing and being replaced by first-year or young sea ice. Young ice melts more rapidly than multi-year ice, which could negatively affect caribou movements between islands and between islands and the mainland (Paquette 2020). Recent studies of sea-ice break-up around Banks Island suggest break-up will occur 2–3 days earlier for each 1 °C increase in temperature (Cooley et al. 2020). The projected changes in sea ice could severely hinder Peary caribou movement between islands in the future if animals are unable to adapt to changing conditions.
Peary caribou around Banks Island and Victoria Island generally move north in the spring to their calving areas as the snow conditions change and forage becomes accessible (Olohaktomiut HTC 2013; Paulatuk HTC 2013; Sachs Harbour HTC 2013). In the fall, the animals move back south to their wintering areas (Olohaktomiut HTC 2013; Paulatuk HTC 2013; Sachs Harbour HTC 2013). Caribou movement around King Williams Island, Nunavut, occurs over several months in spring (April–June) and fall (September–November; Ljubicic et al. 2017). Unfortunately, the months and duration of the spring and fall movements in our study area were not discussed in community workshops (Johnson et al. 2016). However, the broad seasonal categories defined by Johnson et al. (2016) included months outside the spring and fall movement periods. The inclusion of months prior to fall sea-ice freeze-up in the summer–fall season (July–October) would have under-estimated average sea-ice concentrations used by caribou in early fall, whereas late fall sea-ice concentrations would have overestimated by the inclusion of winter months (November–March). Whilst this likely introduced additional variability in our analysis, we were still able to observe differences in young sea ice across the three zones around Banks Island that are consistent with Indigenous Knowledge of caribou movement behaviour in the area.
Indigenous Knowledge indicates that caribou have and continue to use the crossing between Banks and Victoria islands (Zone 1; ECCC 2021; Johnson et al. 2016). The increases in young sea ice suggest that conditions remain favourable for caribou movement, at least in spring when caribou are moving north to access suitable foraging locations. The decreases observed in summer–fall sea ice since 2000 could become problematic for caribou as they move south across the sea ice to their wintering areas. These changes could indicate later fall freeze-up (Paquette 2020). However, Indigenous Knowledge suggests that multiple factors contribute to sea-ice conditions, including distance from land (Paquette 2020). The relative short distance between shorelines in Zone 1 (15–20 km) may maintain favourable conditions for caribou, depending on when they move across the sea ice in the fall.
Indigenous Knowledge indicates that caribou used to cross Zone 2 between Banks Island and the mainland a few decades ago. Our results suggest that young sea-ice concentration in the zone decreased after 2000. Decreases in spring sea-ice cover could delay or prevent movement from wintering areas on the mainland to areas used in spring on Banks Island for calving with highly nutritious and digestible forage that is essential for calf-provisioning (Paulatuk HTC 2013; Sachs Harbour HTC 2013; Le Corre et al. 2017; Gurarie et al. 2019). Similarly, decreases in fall sea-ice concentrations could create barriers preventing caribou from moving back south to access areas with sufficient resources to increase their fat reserves for winter. These delays could result in malnutrition or death. In 2010, several caribou were found frozen after they fell through the ice and drowned near Victoria Island (Paquette 2020). Caribou from the Dolphin and Union herd have also been found frozen on the ice and islands near an open-water area where they used to cross (Dumond & Lee 2013). The general decline in extent, thickness and duration and delay in freeze-up of sea ice over the last decades (Panikkar et al. 2018) likely have contributed to why caribou have stopped crossing in Zone 2.
Finally, the zone between Banks and Melville islands (Zone 3) appeared to experience relatively constant young sea-ice concentrations in spring and winter. Temporal changes in conditions were most pronounced in the summer–fall (sea-ice freeze-up) with fewer positive anomalies occurring after 2000. Whilst caribou tracks have been observed across Zone 3 (Olohaktomiut HTC 2016), more work is needed to understand the frequency, direction and environmental conditions that might influence sea-ice crossings in the area. Our MaxEnt results suggest that the occurrence of Peary caribou on Melville Island is restricted in winter. The deterioration of fall sea-ice conditions in Zone 3 could prevent caribou from escaping potentially unfavourable conditions on Melville Island by moving to Banks Island in the fall. More data on the timing of caribou crossings in this zone and winter data on Peary caribou locations for Banks Island would be needed to evaluate this hypothesis.
Our ability to detect statistical trends was limited because of the high variability in annual ice concentrations. In addition to the inclusion of months outside the timing of caribou movement (see above), errors in the identification of sea-ice types (Couture 2022) may have contributed to the variability. Alternatively, the high variability could be linked to El Niño years, which result in a decrease, followed by non-El Niño years, which are usually associated with an increase in young ice (Tivy et al. 2011). Regardless, caribou will find it harder to cross sea ice because of accelerated warming (Cooley et al. 2020) and a sustained decline of sea-ice extent (i.e., –54 000 km²/year; Yadav et al. 2020) associated with climate change. Earlier sea-ice break-up (Howell et al. 2009) that encourages increased maritime traffic (Paquette 2020) could further impact crossing areas that are currently being used by caribou (Johnson et al. 2016; Carter et al. 2017). The use of other indicators of sea-ice conditions for caribou, such as timing of sea-ice break-up and freeze-up (see Paquette 2020), could help reduce uncertainty of temporal trends by providing a more complete picture of changing sea-ice conditions across the three zones.
Snow simulation and caribou
Our results highlight the spatiotemporal variability in snow density across our study area. The location and amount of area with dense snow, creating unfavourable foraging conditions for caribou on Banks Island and the Melville Island Complex, differed across the seasons. The seasonal changes in snow conditions, in turn, influenced the distribution of Peary caribou across our study areas, with caribou occupying areas where the snow was less dense and forage was likely accessible (Ouellet et al. 2017).
The meteorological forcing data are the primary source of uncertainty in our snow models (Ouellet et al. 2017). However, Martineau et al. (2022) demonstrated the efficiency of OSSA in simulating slab density, which is central to the calculation of CT350, by comparing OSSA snow outputs to in situ field measurements. This work provides assurance that, despite this source of uncertainty, our results are reliable. Averaging snow properties over multiple years may have also created a temporal mismatch between when caribou occupied an area and the snow conditions, in some instances; caribou may have occupied areas where average snow density was high only in years when snow was less dense. A logical next step would be to analyse these results on an annual time step to see if increased temporal matching between Peary caribou observations and snow conditions improves the ability of models to predict Peary caribou occurrence (Boyce et al. 2002; Martineau et al. 2022).
Distribution modelling (MaxEnt probabilistic model)
Previous studies have highlighted the influence of vegetation on changes in the distribution of Peary caribou across seasons (e.g., Tews et al. 2007). Our study highlights the additional importance of snow in determining the accessibility of food. The majority of our top models included both cumulative snow density and vegetation with seasonal difference in their relative importance. Notably, the explanatory power of vegetation for Banks Island and Melville Island was highest for summer–fall, when cumulative snow densities were at their lowest (<800 cm). The lack of data on caribou locations on Banks Island makes it difficult to draw general inferences about winter. Nevertheless, the relative importance of snow on Melville Island increased from spring to winter as caribou occupied areas when higher snow densities likely limited forage accessibility.
Results by Vikhamar-Schuler et al. (2013) and Ouellet et al. (2017) suggest that cumulative snow thickness exceeding 350 kg/m³ could be problematic for foraging reindeer and caribou. Ouellet et al. (2017) showed that caribou numbers tended to decline with snow thickness of high density over ca. 2500 cm. Despite the overall decrease in the probability of occurrence with increasing snow density, the probability of caribou presence remained high at snow densities of 2000 to 7000 cm (more details in the Supplementary material). This suggests that many caribou may have experienced suboptimal foraging conditions during our study. Peary caribou numbers on Banks Island and the surrounding areas declined sharply from the 1980s to the late 1990s because of several successive severe winter weather events (Johnson et al. 2016). Although Peary caribou numbers were no longer declining, the persistence of suboptimal foraging conditions in the area could explain why Peary caribou numbers remained low during 2000–2013.
Conclusion
In this paper, we calculated and analysed young sea-ice anomalies over 1983–2019 in specific areas around Banks Island to evaluate the impact of the anomalies on caribou movement across sea ice. More specifically, we examined whether changes in young sea-ice concentration were consistent with observations of caribou movement described by Indigenous Knowledge holders. Similar to other studies (Paquette 2020), our analysis revealed high inter-annual and seasonal variability, making it difficult to detect spatial and temporal trends. Nevertheless, we did detect reductions in spring sea-ice concentrations relative to the 40-yr average in Zone 2, especially after 2000, which may explain why caribou are no longer observed moving between Banks Island and the mainland (Paulatuk HTC 2013). Caribou moving from Banks Island to mainland in the fall would remain trapped on the mainland, unable to move north the following spring. These results suggest that characterizing spatial and temporal changes in sea-ice anomalies may be useful for monitoring and mitigating the impacts of climate change on caribou movement between islands and the mainland in the Arctic (ECCC 2021). For example, declines in young sea ice in the fall could be used as a warning sign to limit shipping traffic late in the season, as a mitigation strategy to prevent additional delays in the timing of young sea-ice freeze-up that would hinder caribou movement. A more fulsome evaluation of the utility of sea-ice anomalies would require not only additional work to refine the timing and duration of spring and fall movement in our study area but also studies like ours in different areas across the Canadian Arctic Archipelago.
Our adjustments and refinements to snow simulations with the OSSA (Ouellet et al. 2017) proved useful in predicting spatial variation in the distribution of Peary caribou in different seasons. Our results are consistent with the prevailing notion that food limitation is the principal factor driving changes in the distribution and abundance of Peary caribou. Whilst vegetation may determine forage availability, snow or lack thereof will determine forage accessibility. As such, future studies forecasting changes in snow condition will be essential to the development of climate smart strategies for the species’ recovery, including identifying areas for habitat protection that contribute to species persistence over the short and longer term (ECCC 2021).
Acknowledgements
The authors are extremely grateful to the Inuvialuit and Inuit communities who shared their knowledge, stories and love of Peary caribou and the connection between caribou, people and the Arctic landscape. See Johnson et al. (2016) for all the collaborators. Specifically, the authors thank the following: Resolute Bay Hunters and Trappers Organization, Philip Manik Sr., Community of Resolute Bay, NU; Iviq Hunters and Trappers Association, Amon Akeeagok, Jaypetee Akeeagok, Charlie Noah, Community of Grise Fiord, NU; Gjoa Haven Hunters and Trappers Association, James Qitsualik Taqaugak, Community of Gjoa Haven, NU; Kurairojuark Hunters and Trappers Association, John Kayasark, Zachary Oogark, Columban Pujuardjok, Community of Kugaaruk, NU; Spence Bay Hunters & Trappers Association, Abel Aqqaq, Peter Qayutinuak Sr., Community of Taloyoak, NU; Ekaluktutiak Hunters and Trappers Organization, Howard Greenley, James Panioyak, George Angohiatok, Jimmy Haniliak, Community of Cambridge Bay, NU; Sachs Harbour Hunters and Trappers Committee, Issiac Elanik, Vernon Amos, John Lucus Jr., Joey Carpenter, Community of Sachs Harbour, NWT; Olohaktomiut Hunters & Trappers Committee, Bradley Carpenter, Joshua Oliktoak, Community of Ulukhaktok, NWT; Paulatuk Hunters and Trappers Committee, Raymond Ruben Sr., Community of Paulatuk, NWT. The authors thank Mael Le Corre and Bernardo Brandao Niebuhr for their helpful comments on earlier versions of the manuscript. The authors are also grateful to Isabelle Duclos and Carine Côté-Germain for producing Fig. 1.
References
| Ahmed S.E., McInerny G., O’Hara K., Harper R., Salido L., Emmott S. & Joppa L.N. 2015. Scientists and software—surveying the species distribution modelling community. Diversity and Distributions 21, 258–267, doi: 10.1111/ddi.12305. |
| Bartelt P. & Lehning M. 2002. A physical SNOWPACK model for the Swiss avalanche warning. Part I: numerical model. Cold Regions Science and Technology 35, 123–145, doi: 10.1016/S0165-232X(02)00074-5. |
| Beaudoin-Galaise M. 2016. Amélioration de la résolution spatiale de simulations de neige du modèle SNOWPACK dans un contexte de l’accès à la nourriture du caribou de Peary. (Improving the spatial resolution of snow simulations from the SNOWPACK model in the context of Peary caribou food access.) MSc thesis, Université de Sherbrooke, Sherbrooke, QC. |
| Blanchard-Wrigglesworth E., Armour K.C., Bitz C.M. & DeWeaver E. 2011. Persistence and inherent predictability of Arctic sea ice in a GCM ensemble and observations. Journal of Climate 24, 231–250, doi: 10.1175/2010JCLI3775.1. |
| Boelman N.T., Liston G.E., Gurarie E., Meddens A.J.H., Mahoney P.J., Kirchner P.B., Bohrer G., Brinkman T.J., Cosgrove C.L., Eitel J.U.H., Hebblewhite M., Kimball J.S., LaPoint S., Nolin A.W., Pedersen S.H., Prugh L.R., Reinking A.K. & Vierling L.A. 2019. Integrating snow science and wildlife ecology in Arctic–boreal North America. Environmental Research Letters 14, 010401, doi: 10.1088/1748-9326/aaeec1. |
| Bokhorst S., Pedersen S.H., Brucker L., Anisimov O., Bjerke J.W., Brown R.D., Ehrich D., Essery R L.H., Heilig A., Ingvander S., Johansson C., Johansson M., Jónsdóttir I.S., Inga N., Luojus K., Macelloni G., Mariash H., McLennan D., Rosqvist G.N., Sato A., Savela H., Schneebeli M., Sokolov A., Sokratov S.A., Terzago S., Vikhamar-Schuler D., Williamson S., Qiu Y. & Callaghan T.V. 2016. Changing Arctic snow cover: a review of recent developments and assessment of future needs for observations, modelling, and impacts. Ambio 45, 516–537, doi: 10.1007/s13280-016-0770-0. |
| Boyce M., Vernier P.R., Nielsen S.E. & Schmiegelow F.K.A. 2002. Evaluating resource selection functions. Ecological Modelling 157, 281–300, doi: 10.1016/S0304-3800(02)00200-4. |
| Bushuk M. & Giannakis D. 2017. The seasonality and interannual variability of Arctic sea ice reemergence. Journal of Climate 30, 4657–4676, doi: 10.1175/JCLI-D-16-0549.1. |
| Cai Q., Wang J., Beletsky D., Overland J.E., Ikeda M. & Wan L. 2021. Summer Arctic sea ice decline during 1850–2017 and the amplified Arctic warming during the recent decades. Environmental Research Letters 16, 034015, doi: 10.1088/1748-9326/abdb5f. |
| Callaghan T.V., Johansson M., Brown R.D., Groisman P.Y., Labba N., Radionov V., Barry R.G., Bulygina O.N., Essery R.L.H., Frolov D.M., Golubev V.N., Grenfell T.C., Petrushina M.N., Razuvaev V.N., Robinson D.A., Romanov P., Shindell D., Shmakin A.B., Sokratov S.A., Warren S. & Yang D. 2011. The changing face of Arctic snow cover: a synthesis of observed and projected changes. Ambio 40, 17–31, doi: 10.1007/s13280-011-0212-y. |
| Canadian Ice Service 2005. MANICE: manual of standard procedures for observing and reporting ice conditions. Accessed on the internet at https://www.canada.ca/en/environment-climate-change/services/weather-manuals-documentation/manice-manual-of-ice.html on May 2021. |
| Carter N., Dawson J., Joyce J. & Ogilvie A. 2017. Arctic corridors and Northern voices: governing marine transportation in the Canadian Arctic. Gjoa Haven, Nunavut. Ottawa: University of Ottawa. |
| CAVM Team 2003. Circumpolar Arctic vegetation Map. Scale: 1:7,500,000. Conservation of Arctic Flora and Fauna (CAFF) Map no. 1. Anchorage: US Fish and Wildlife Service. |
| Cooley S.W., Ryan J.C., Smith L.C., Horvat C., Pearson B., Dale B. & Lynch A.H. 2020. Coldest Canadian Arctic communities face greatest reductions in shorefast sea ice. Nature Climate Change 10, 533–538, doi: 10.1038/s41558-020-0757-5. |
| COSEWIC (Committee on the Status of Endangered Wildlife in Canada) 2015. COSEWIC assessment and status report on the Peary caribou Rangifer tarandus pearyi in Canada. Ottawa: Committee on the Status of Endangered Wildlife in Canada. |
| Couture G. 2022. Analyses spatiotemporelles des conditions de glace de mer et des tendances de formation des polynies de l’Archipel Arctique Canadien. (Spatiotemporal analyses of sea-ice conditions and polynya formation trends in the Canadian Arctic Archipelago.) MSc thesis, Université de Sherbrooke, Sherbrooke, QC. |
| Derksen C. & Brown R. 2012. Spring snow cover extent reductions in the 2008–2012 period exceeding climate model projections. Geophysical Research Letters 39, L19504, doi: 10.1029/2012GL053387. |
| Derksen C., Burgess D., Duguay C., Howell S., Mudryk L., Smith S., Thackeray C. & Kirchmeier-Young M. 2019. Changes in snow, ice, and permafrost across Canada. In E. Bush & D.S. Lemmen (eds.): Canada’s changing climate report. Pp. 194–260. Ottawa: Government of Canada. |
| Dolant C., Langlois A., Brucker L., Royer A., Roy A. & Montpetit B. 2017. Meteorological inventory of rain-on-snow events in the Canadian Arctic Archipelago and satellite detection assessment using passive microwave data. Physical Geography 39, 428–444, doi: 10.1080/02723646.2017.1400339. |
| Dolant C., Langlois A., Montpetit B., Brucker L., Roy A. & Royer A. 2016. Development of a rain‐on‐snow detection algorithm using passive microwave radiometry. Hydrological Processes 30, 3184–3196, doi: 10.1002/hyp.10828. |
| Dolant C., Montpetit B., Langlois A., Brucker L., Zolina O., Johnson C.A., Royer A. & Smith P. 2018. Assessment of the barren ground caribou die-off during winter 2015–2016 using passive microwave observations. Geophysical Research Letters 45, 4908–4916, doi: 10.1029/2017GL076752. |
| Dumond M. & Lee D.S. 2013. Dolphin and Union caribou herd status and trend. Arctic 66, 329–337, doi: 10.14430/Arctic4311. |
| ECCC (Environment and Climate Change Canada) 2021. Recovery strategy for the Peary Caribou (Rangifer tarandus pearyi) in Canada [Proposed]. Species at risk act recovery strategy series. Ottawa: Environment and Climate Change Canada. |
| Elith J., Graham C., Anderson R., Dudik M., Ferrier S., Guisan A., Hijmans R., Huettmann F., Leathwick J., Lehmann A., Li J., Lohmann L., Loiselle B., Manion G., Moritz C., Nakamura M., Nakazawa Y., Overton J., Peterson A., Phillips S., Richardson K., Scachetti-Pereira R., Schapire R., Soberon J., Williams S., Wisz M. & Zimmermann N. 2006. Novel methods improve prediction of species’ distributions from occurrence data. Ecography 29, 129–151, doi: 10.1111/j.2006.0906-7590.04596.x. |
| Elith J. & Leathwick J.R. 2009. Species distribution models: ecological explanation and prediction across space and time. Annual Review of Ecology, Evolution, and Systematics 40, 677–697, doi: 10.1146/annurev.ecolsys.110308.120159. |
| Elith J., Phillips S.J., Hastie T., Dudík M., Chee Y.E. & Yates C.J. 2011. A statistical explanation of MaxEnt for ecologists. Diversity and Distributions 17, 43–57, doi: 10.1111/j.1472-4642.2010.00725.x. |
| Forbes B.C., Kumpula T., Meschtyb N., Laptander R., MacIas-Fauria M., Zetterberg P., Verdonen M., Skarin A., Kim K.Y., Boisvert L.N., Stroeve J.C. & Bartsch, A. 2016. Sea ice, rain-on-snow and tundra reindeer nomadism in Arctic Russia. Biology Letters 12, article no. 20160466, doi: 10.1098/rsbl.2016.0466. |
| Gardner A., Moholdt G., Arendt A. & Wouters B. 2012. Accelerated contributions of Canada’s Baffin and Bylot Island glaciers to sea level rise over the past half century. The Cryosphere 6, 1103–1125, doi: 10.5194/tc-6-1103-2012. |
| Gouttevin I., Langer M., Lowe H., Boike J., Proksch M. & Schneebeli M. 2018. Observation and modelling of snow at a polygonal tundra permafrost site: spatial variability and thermal implications. The Cryosphere 12, 3693–3717, doi: 10.5194/tc-12-3693-2018. |
| Government of Canada 2020a. Canadian digital elevation model, 1945–2011. Accessed on the internet at https://open.canada.ca/data/en/dataset/7f245e4d-76c2-4caa-951a-45d1d2051333 on January 2021. |
| Government of Canada 2020b. Canadian land cover, circa 2000 (vector). GeoBase Series 1996–2005. Accessed on the internet at https://open.canada.ca/data/en/dataset/97126362-5a85-4fe0-9dc2-915464cfdbb7 on January 2021. |
| Government of Canada 2020c. Weekly regional ice charts—black and white. Accessed on the internet at https://www.canada.ca/en/environment-climate-change/services/ice-forecasts-observations/latest-conditions/archive-overview.html on January 2021. |
| Groot Zwaaftink C.D., Cagnati A., Crepaz A., Fierz C., Macelloni G., Valt M. & Lehning M. 2013. Event-driven deposition of snow on the Antarctic Plateau: analyzing field measurements with SNOWPACK. The Cryosphere 7, 333–347, doi: 10.5194/tc-7-333-2013. |
| Guarino M.V., Sime L.C., Schröeder D., Malmierca-Vallet I., Rosenblum E., Ringer M., Ridley J., Feltham D., Bitz C., Steig E.J., Wolff E., Stroeve J. & Sellar A. 2020. Sea-ice-free Arctic during the last interglacial supports fast future loss. Nature Climate Change 10, 928–932, doi: 10.1038/s41558-020-0865-2. |
| Gurarie E., Hebblewhite M., Joly K., Kelly A.P., Adamczewski J., Davidson S.C., Davison T., Gunn A., Suitor M.J., Fagan W.F. & Boelman N. 2019. Tactical departures and strategic arrivals: divergent effects of climate and weather on caribou spring migrations. Ecosphere 10, e02971, doi: 10.1002/ecs2.2971. |
| Hansen B.B., Isaksen K., Benestad R.E., Kohler J., Pedersen Å., Loe L.E., Coulson S.J., Larsen J.O. & Varpe Ø. 2014. Warmer and wetter winters: characteristics and implications of an extreme weather event in the High Arctic. Environmental Research Letters 9, article no. 114021, doi: 10.1088/1748-9326/9/11/114021. |
| Holland M.M., Serreze M.C. & Stroeve J. 2010. The sea ice mass budget of the Arctic and its future change as simulated by coupled climate models. Climate Dynamics 34, 185–200, doi: 10.1007/s00382-008-0493-4. |
| Howell S.E., Duguay C.R. & Markus T. 2009. Sea ice conditions and melt season duration variability within the Canadian Arctic Archipelago: 1979–2008. Geophysical Research Letters 36, L1050, doi: 10.1029/2009GL037681. |
| Il Jeong D. & Sushama L. 2018. Rain-on-snow events over North America based on two Canadian regional climate models. Climate Dynamics 50, 303–316, doi: 10.1007/s00382-017-3609-x. |
| Jenkins D.A., Lecomte N., Andrews G., Yannic G. & Schaefer J.A. 2020. Biotic interactions govern the distribution of coexisting ungulates in the Arctic Archipelago—a case for conservation planning. Global Ecology and Conservation 24, e01239, doi: 10.1016/j.gecco.2020.e01239. |
| Jenkins D.A., Lecomte N., Schaefer J.A., Olsen S.M., Swingedouw D., Côté S.D., Pellissier L. & Yannic G. 2016. Loss of connectivity among island-dwelling Peary caribou following sea ice decline. Biology Letters 12, article no. 20160235, doi: 10.1098/rsbl.2016.0235. |
| Johnson C.A., Neave E., Blukacz-Richards A., Banks S.N. & Quesnelle P.E. 2016. Knowledge assessment (community and scientific) to inform the identification of critical habitat for Peary caribou, Rangifer tarandus pearyi, in the Canadian Arctic. Ottawa: Environment and Climate Change Canada. |
| Joint Technical Commission for Oceanography and Marine Meteorology 2014. Sea ice nomenclature. Accessed on the internet at https://library.wmo.int/doc_num.php?explnum_id=4651 on 20 July 2022. |
| Kaluskar S., Johnson C.A., Blukacz-Richards E.A., Ouellet F., Kim D.K. & Arhonditsis G. 2020. A stochastic modelling framework to accommodate the inter-annual variability of habitat conditions for Peary caribou (Rangifer tarandus pearyi) populations. Ecological Informatics 56, article no. 101013, doi: 10.1016/j.ecoinf.2019.101013. |
| Kohler J. & Aanes R. 2004. Effect of winter snow and ground-icing on a Svalbard reindeer population: results of a simple snowpack model. Arctic, Antarctic, and Alpine Research 36, 333–341, doi: 10.1657/1523-0430(2004)036[0333:EOWSAG]2.0.CO;2. |
| Langlois A., Johnson C.A., Montpetit B., Royer A., Blukacz-Richards E.A., Neave E., Dolant C., Roy A., Arhonditsis G., Kim D.K., Kaluskar S. & Brucker L. 2017. Detection of rain-on-snow (ROS) events and ice layer formation using passive microwave radiometry: a context for Peary caribou habitat in the Canadian Arctic. Remote Sensing of Environment 189, 84–95, doi: 10.1016/j.rse.2016.11.006. |
| Le Corre M., Dussault C. & Côté S.D. 2017. Weather conditions and variation in timing of spring and fall migrations of migratory caribou. Journal of Mammalogy 98, 260–271, doi: 10.1093/jmammal/gyw177. |
| Lehning M., Bartelt P., Brown B., Fierz C. & Satyawali P. 2002. A physical SNOWPACK model for the Swiss avalanche warning. Part II. Snow microstructure. Cold Regions Science and Technology 35, 147–167, doi: 10.1016/S0165-232X(02)00073-3. |
| Liston G.E. & Hiemstra C.A. 2011. The changing cryosphere: pan-Arctic snow trends (1979–2009). Journal of Climate 24, 5691–5712, doi: 10.1175/JCLI-D-11-00081.1. |
| Ljubicic G., Oberndorfer E. & Smith G. 2017. The curious case of King William Island, Nunavut: an island overlooked in caribou research. Arctic 70, 107–117, doi: 10.14430/arctic4638. |
| Ljubicic G., Okpakok S., Robertson S. & Mearns R. 2018. Uqsuqtuurmiut inuita tuktumi qaujimaningit (Inuit knowledge of caribou from Gjoa Haven, Nunavut): collaborative research contributions to co-management efforts. Polar Record 54, 213–233, doi: 10.1017/S0032247418000372. |
| Martineau C., Langlois A., Gouttevin I., Royer A., Neave E. & Johnson C.A. 2022. Improving Peary caribou presence prediction from the MaxEnt model using spatialized snow simulations: a case study over Bathurst Island Complex, Nunavut, Canada. Arctic 75, 55–71, doi: 10.14430/arctic74868. |
| Maslanik J., Stroeve J., Fowler C. & Emery W. 2011. Distribution and trends in Arctic sea ice age through spring 2011. Geophysical Research Letters 38, L13502, doi: 10.1029/2011GL047735. |
| Meloche J., Royer A., Langlois A., Rutter N. & Sasseville V. 2021. Improvement of microwave emissivity parameterization of frozen Arctic soils using roughness measurements derived from photogrammetry. International Journal of Digital Earth 14, 1380–1396, doi: 10.1080/17538947.2020.1836049. |
| Mudryk L., Derksen C., Howell S., Laliberté F., Thackeray C., Sospedra-Alfonso R., Vionnet V., Kushner P. & Brown R. 2018. Canadian snow and sea ice: historical trends and projections. The Cryosphere 12, 1157–1176, doi: 10.5194/tc-12-1157-2018. |
| NOAA (National Oceanic and Atmospheric Administration) 2020. North American Regional Reanalysis (NARR) for Arctic research. Dataset. Physical Sciences Laboratory, NOAA. Accessed on the internet at https://psl.noaa.gov/data/narr/ on January 2021. |
| Olohaktomiut HTC (Hunters and Trappers Committee) 2013. Summary of HTC and public Peary caribou federal recovery strategy development community technical meetings—March 4, 2013. Unpublished report. Ulukhaktok, NT: Canadian Wildlife Service. |
| Olohaktomiut HTC (Hunters and Trappers Committee) 2016. Summary of HTC and public meeting for the draft Peary caribou recovery strategy—March 9, 2016. Unpublished report. Ulukhaktok, NT: Canadian Wildlife Service. |
| Ouellet F., Langlois A., Blukacz-Richards E.A., Johnson C.A., Royer A., Neave E. & Larter N.C. 2017. Spatialization of the SNOWPACK snow model for the Canadian Arctic to assess Peary caribou winter grazing conditions. Physical Geography 38, 143–158, doi: 10.1080/02723646.2016.1274200. |
| Panikkar B., Lemmond B., Else B. & Murray M. 2018. Ice over troubled waters: navigating the Northwest Passage using Inuit knowledge and scientific information. Climate Research 75, 81–94, doi: 10.3354/cr01501. |
| Papasodoro C., Berthier E., Royer A., Zdanowicz C. & Langlois A. 2015. Area, elevation and mass changes of the two southernmost ice caps of the Canadian Arctic Archipelago between 1952 and 2014. The Cryosphere 9, 1535–1550, doi: 10.5194/tc-9-1535-2015. |
| Paquette E. 2020. Potential impacts of sea ice and ship traffic change to caribou sea ice crossing areas surrounding King William Island, Nunavut, Canada. MSc thesis, Department of Geography, Carleton University, Ottawa. |
| Parker G.R., Thomas D.C., Broughton E. & Gray D.R. 1975. Crashes of muskox and Peary caribou populations in 1973–74 on the Parry Islands, Arctic Canada. Progress Notes 56. Ottawa: Canadian Wildlife Service, Environment Canada. |
| Paulatuk HTC (Hunters and Trappers Committee). 2013. Summary of HTC and public Peary caribou federal recovery strategy development community technical meeting—March 6, 2013. Unpublished report. Paulatuk, NT: Canadian Wildlife Service. |
| Phillips S.J., Anderson R.P. & Schapire R.E. 2006. Maximum entropy modeling of species geographic distributions. Ecological Modelling 190, 231–259, doi: 10.1016/j.ecolmodel.2005.03.026. |
| Phillips S.J. & Dudík M. 2008. Modeling of species distributions with MaxEnt: new extensions and a comprehensive evaluation. Ecography 31, 161–175, doi: 10.1111/j.0906-7590.2008.5203.x. |
| Phillips S.J., Dudík M. & Schapire R.E. 2019. Maxent software for modeling species niches and distributions. Version 3.4.1. Accessed on the internet at http://biodiversityinformatics.amnh.org/open_source/maxent/ on January 2021. |
| Portet S. 2020. A primer on model selection using the Akaike Information Criterion. Infectious Disease Modelling 5, 111–128, doi: 10.1016/j.idm.2019.12.010. |
| Rigor I.G. & Wallace J.M. 2004. Variations in the age of Arctic sea‐ice and summer sea‐ice extent. Geophysical Research Letters 31, L09401, doi: 10.1029/2004GL019492. |
| Rinke A. & Dethloff K. 2008. Simulated circum-Arctic climate changes by the end of the 21st century. Global and Planetary Change 62, 173–186, doi: 10.1016/j.gloplacha.2008.01.004. |
| Royer A., Dominé F., Roy A., Langlois A., Marchand N. & Davesne G. 2021. New northern snowpack classification linked to vegetation cover on a latitudinal mega-transect across northeastern Canada. Écoscience 28, 225–242, doi: 10.1080/11956860.2021.1898775. |
| Sachs Harbour HTC (Hunters and Trappers Committee) 2013. Summary of HTC, elder and public Peary caribou federal recovery strategy development community technical meetings—March 5, 2013. Unpublished report. Sachs Harbour, NT: Canadian Wildlife Service. |
| Sachs Harbour HTC (Hunters and Trappers Committee) 2016. Summary of HTC and public meetings for the draft Peary caribou recovery strategy—March 8, 2016. Unpublished report. Sachs Harbour, NT: Canadian Wildlife Service. |
| Serreze M.C. & Barry R.G. 2011. Processes and impacts of Arctic amplification: a research synthesis. Global and Planetary Change 77, 85–96, doi: 10.1016/j.gloplacha.2011.03.004. |
| Serreze M.C. & Francis J.A. 2006. The Arctic amplification debate. Climatic Change 76, 241–264, doi: 10.1007/s10584-005-9017-y. |
| Serreze M.C. & Stroeve J. 2015. Arctic sea ice trends, variability and implications for seasonal ice forecasting. Philosophical Transactions of the Royal Society A 373, article no. 20140159, doi: 10.1098/rsta.2014.0159. |
| Smith A.B. & Santos M.J. 2020. Testing the ability of species distribution models to infer variable importance. Ecography 43, 1801–1813, doi: 10.1111/ecog.05317. |
| Sobota I., Weckwerth P. & Grajewski T. 2020. Rain-on-snow (ROS) events and their relations to snowpack and ice layer changes on small glaciers in Svalbard, the High Arctic. Journal of Hydrology 590, article no. 125279, doi: 10.1016/j.jhydrol.2020.125279. |
| Stien A., Loel L.E., Mysterud A., Severinsen T., Kohler J. & Langvatn R. 2010. Icing events trigger range displacement in a High-Arctic ungulate. Ecology 91, 915–920, doi: 10.1890/09-0056.1. |
| Stocker T.F., Qin D., Plattner G.K., Tignor M., Allen S.K., Boschung J., Nauels A., Xia Y., Bex B. & Midgley B. (eds.) 2013. Climate change 2013: the physical science basis. Contribution of Working Group I to the fifth assessment report of the Intergovernmental Panel on Climate Change. Cambridge: Cambridge University Press. |
| Stroeve J., Holland M.M., Meier W., Scambos T. & Serreze M. 2007. Arctic sea ice decline: faster than forecast. Geophysical Research Letters 34, L9501, doi: 10.1029/2007GL029703. |
| Stroeve J. & Notz D. 2018. Changing state of Arctic sea ice across all seasons. Environmental Research Letters 13, article no. 103001, doi: 10.1088/1748-9326/aade56. |
| Tews J., Ferguson M.A.D. & Fahrig L. 2007. Potential net effects of climate change on High Arctic Peary caribou: lessons from a spatially explicit simulation model. Ecological Modelling 207, 85–89, doi: 10.1016/j.ecolmodel.2007.04.011. |
| Tivy A., Howell S.E.L., Alt B., McCourt S., Chagnon R., Crocker G., Carrieres T. & Yackel J.J. 2011. Trends and variability in summer sea ice cover in the Canadian Arctic based on the Canadian Ice Service Digital Archive, 1960–2008 and 1968–2008. Journal of Geophysical Research—Oceans 116, C03007, doi: 10.1029/2009JC005855. |
| Tomaselli M., Kutz S., Gerlach C. & Checkley S. 2018. Local knowledge to enhance wildlife population health surveillance: conserving muskoxen and caribou in the Canadian Arctic. Biological Conservation 217, 337–348, doi: 10.1016/j.biocon.2017.11.010. |
| Turunen M., Soppela P., Kinnunen H., Sutinen M.L. & Martz F. 2009. Does climate change influence the availability and quality of reindeer forage plants? Polar Biology 32, 813–832, doi: 10.1007/s00300-009-0609-2. |
| Tyler N.J.C. 2010. Climate, snow, ice, crashes, and declines in populations of reindeer and caribou (Rangifer tarandus L.). Ecological Monographs 80, 197–219, doi: 10.1890/09-1070.1. |
| Uboni A., Horstkotte T., Kaarlejärvi E., Sévêque A., Stammler F., Olofsson J., Forbes B.C. & Moen J. 2016. Long-term trends and role of climate in the population dynamics of Eurasian reindeer. PLoS One 11, e0158359, doi: 10.1371/journal.pone.0158359. |
| Vikhamar-Schuler D., Hanssen-Bauer I., Schuler T.V., Mathiesen S.D. & Lehning M. 2013. Use of a multilayer snow model to assess grazing conditions for reindeer. Annals of Glaciology 54, 214–226, doi: 10.3189/2013AoG62A306. |
| Vincent L.A., Zhang X., Mekis E., Wan H. & Bush E.J. 2018. Changes in Canada’s climate: trends in indices based on daily temperature and precipitation data. Atmosphere—Ocean 56, 332–349, doi: 10.1080/07055900.2018.1514579. |
| Yadav J., Kumar A. & Mohan R. 2020. Dramatic decline of Arctic sea ice linked to global warming. Natural Hazards 103, 2617–2621, doi: 10.1007/s11069-020-04064-y. |
Related Articles
Louise Chawla
Hanna К. Lappalainen
Tuukka Petäjä
Joni Kujansuu
Veli-Matti Kerminen
et al.
Ahmet Kılınç
Martin Stanisstreet
Edward Boyes
Fiona Taber
Neil Taylor
Georgia Liarakou
Ilias Athanasiadis
Costas Gavrilakis